NKG2D
NKG2D is an activating receptor (transmembrane protein) belonging to the NKG2 family of C-type lectin-like receptors.[3] NKG2D is encoded by KLRK1 (killer cell lectin like receptor K1) gene which is located in the NK-gene complex (NKC) situated on chromosome 6 in mice[4] and chromosome 12 in humans.[5] In mice, it is expressed by NK cells, NK1.1+ T cells, γδ T cells, activated CD8+ αβ T cells and activated macrophages.[6] In humans, it is expressed by NK cells, γδ T cells and CD8+ αβ T cells.[7] NKG2D recognizes induced-self proteins from MIC and RAET1/ULBP families which appear on the surface of stressed, malignant transformed, and infected cells.[8]
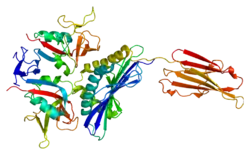


Structure
Human NKG2D receptor complex assembles into a hexameric structure. NKG2D itself forms a homodimer whose ectodomains serve for ligand binding.[9] Each NKG2D monomer is associated with DAP10 dimer. This association is maintained by ionic interaction of a positively charged arginine present in a transmembrane segment of NKG2D and negatively charged aspartic acids within both transmembrane regions of DAP10 dimer.[10] DAP10 functions as an adaptor protein and transduces the signal after the ligand binding by recruiting the p85 subunit of PI3K and Grb2-Vav1 complex which are responsible for subsequent downstream events.[11]
In mice, alternative splicing generates two distinct NKG2D isoforms: the long one (NKG2D-L) and the short one (NKG2D-S). NKG2D-L binds DAP10 similarly to human NKG2D. By contrast, NKG2D-S associates with two adaptor proteins: DAP10 and DAP12.[12] DAP10 recruits the p85 subunit of PI3K and a complex of Grb2 and Vav1.[11] DAP12 bears ITAM motif and activates protein tyrosine kinases Syk and Zap70 signalling.[13]
NKG2D ligands
NKG2D ligands are induced-self proteins which are completely absent or present only at low levels on surface of normal cells, but they are overexpressed by infected, transformed, senescent and stressed cells. Their expression is regulated at different stages (transcription, mRNA and protein stabilization, cleavage from the cell surface) by various stress pathways.[14] Among them, one of the most prominent stress pathways is DNA damage response. Genotoxic stress, stalled DNA replication, poorly regulated cell proliferation in tumorigenesis, viral replication or some viral products activate the ATM and ATR kinases. These kinases initiate the DNA damage response pathway which participates in NKG2D ligand upregulation. DNA damage response thus participate in alerting the immune system to the presence of potentially dangerous cells.[15]
All NKG2D ligands are homologous to MHC class I molecules and are divided into two families: MIC and RAET1/ULBP.
MIC family
Human MIC genes are located within the MHC locus and are composed of seven members (MICA-G), of which only MICA and MICB produce functional transcripts. In mice, MIC genes are absent.[16]
RAET1/ULBP family
Among ten known human RAET1/ULBP genes, six encode functional proteins: RAET1E/ULBP4, RAET1G/ULBP5, RAET1H/ULBP2, RAET1/ULBP1, RAET1L/ULBP6, RAET1N/ULBP3. In mice, proteins from orthologous RAET1/ULBP family fall into three subfamilies: Rae-1, H60, and MULT-1.[16] ULBP2 is a stress-induced ligand often found on senescent cells.[17]
NKG2D ligand regulation
NKG2D ligand expression is regulated on multiple levels such as transcriptional, RNA splicing, posttranscriptional and posttranslational. On the transcriptional level, NKG2D ligands can be regulated by transcription factors or regulatory sequences in various molecular pathways. Also, the regulation of NKG2D ligands after cell stress, proliferation signals, infection or oxidative stress is able to activate a DNA damage response (DDR).[18] Ligation of sensor kinases ATM and ATR leads to activation of different checkpoint kinases, such as Chk1 and Chk2,[19] which are important for the induction of MIC, ULBPs or Reat1 genes [15] ). One of the major signals for cell expression of NKG2D is the triggering of DDR along with the induction of senescence program.[15] RNA splicing is another mechanism influencing NKG2D ligand expression. For MICA,[20] ULBP4[21] and ULBP5,[22] alternative splicing isoforms have been shown. However, the molecular mechanisms of this type of regulation are unknown. In posttranscriptional regulation, stabilization of NKG2D ligand mRNA plays a key role. For example AUF1 protein, which mediates RNA degradation, constitutively targets mRNA of NKG2D ligands.[23] Additionally, surface expression levels of NKG2D can be controlled by soluble forms of various protease-mediated cleavages and exosome expression.[24]
Function
NKG2D is a major recognition receptor for the detection and elimination of transformed and infected cells as its ligands are induced during cellular stress, either as a result of infection or genomic stress such as in cancer.[25] In NK cells, NKG2D serves as an activating receptor, which itself is able to trigger cytotoxicity. The function of NKG2D on CD8+ T cells is to send co-stimulatory signals to activate them.[6]
Role in viral infection
Viruses, as intracellular pathogens, can induce the expression of stress ligands for NKG2D. NKG2D is thought to be important in viral control as viruses have adapted mechanisms by which to evade NKG2D responses.[26] For example, cytomegalovirus (CMV) encodes a protein, UL16, which binds to NKG2D ligands ULBP1 and 2 (thus their name "UL16-binding protein") and MICB, which prevents their surface expression.[27]
Role in tumor control
As cancerous cells are "stressed", NKG2D ligands become upregulated, rendering the cell susceptible to NK cell-mediated lysis. However, some tumor cells have acquired the capacity to evade this immune surveillance. They have created the capacity of reducing and eliminating the high volume of NKG2DL present on the cell surface of tumor cells by secreting metalloproteases that cleave these ligands, and therefore they escape from the control of NK cells and their cytotoxic activity. TGF-β allows immune surveillance escape by inhibiting T and NK cell function.[28] Tumor cells that can evade NKG2D responses are thus more likely to propagate.[26][29]
Role in senescent cell removal
As part of the DNA damage response during induction of cellular senescence, cells upregulate the expression of NKG2D ligands that enable NK-mediated killing of senescent cells via the granule exocytosis pathway.[30][31] Specifically, MICA and ULBP2 proteins on senescent cells are recognized by the NKG2D receptor on Natural Killer cells, which is necessary for efficient recognition and elimination of senescent cells.[30]
Interventions to increase senescent cell surface ligands of the Natural Killer cell receptor NKG2D have been proposed as a senolytic therapy to remove senescent cells.[32]
NKG2D signalling
When NKG2D receptor binds to any of its ligands, an activation cascade starts for activating the respective immune cell. NKG2D does not possess any signaling elements within its intracellular domain. NKG2D forms a homodimer and associates with adaptor proteins in its transmembrane domain to a hexameric complex structure and initiate signaling cascades.[33] In both mice and humans, this signalling depends on the association between NKG2D and DAP10 protein forming a complex. Upon ligand engagement, Tyr-X-X-Meth (YXXM) motif within the cytoplasmic domain of DAP10 recruits PI3K and Grb2 to activate NK cell cytotoxicity pathways.[33] In mice, NKG2D associates with DAP 12, instead of DAP 10, and NKG2D-DAP12 complex is involved in IFN-γ production through the Syk and ZAP70 pathway.[28]
Therefore, NKG2D is implicated in NK and other immune cells activation through the PI3K-AKT pathway. This pathway activation depend on two main basis. The first is the plasticity and structural changes of NKG2D (receptor) when binding to its ligands. The last one is the association of DAP10 in the intracellular domain of the receptor and the recruitment of PI3K and Grb.
NKG2D and NK cells
NK cells are a key part of innate immunity, mainly involved in the early cytolytic defense against infections and tumors. NK cell activity is mediated by a variety of cell surface receptors with stimulatory and inhibitory activity. Under normal conditions, NK cells exist in an inactive state, with signaling dominated by inhibitory receptor activation.
NKG2G is a key stimulatory cell surface receptor. A low expression of the receptor is observed already in the early NK cells precursor stages, also the concentration of receptors is increased with maturation of NK cells.[34] In mice, both NKG2D isoforms were detected. During resting state, predominance of long forms of NKG2D is typical, while in activated cells there is a higher number of short forms.[35]
Interaction with IL-15 receptor (IL-15R) is a crucial factor for development, homeostasis and survival of NK cells and NKG2D signaling seems to be similarly critical.[36] Connection between these two pathways is the binding of DAP10, adaptor protein and signal transducer, which associates with IL-15R or NKG2D, respectively.[37] This phenomenon was proven by experiments on mice knocked out in Klrk1 – such mice have higher proliferation rate, faster differentiation and maturation of NK cells, resulting in a misbalance of immature NK cell subpopulations and higher susceptibility to apoptosis of NK cells.[13]
NKG2D is involved in the generation of peripheral tolerance by the effective downregulation of NKG2D ligands, for the prevention from recognition by NK cells. It is supposed to act as a form of obstacle against NK cells' hyper-responsiveness to ligands, without complete education in bone marrow.[38] Tolerance of NK cells is likewise observed during pregnancy, when placenta produces soluble and exosome-bound ligands for NKG2D and accumulates large number of NK cells which prevent recognition of fetus as non-self.[39]
NKG2D and T cells
For priming of T cells, binding of ligand on T cell receptors (TCR), co-stimulation by membrane receptors and cytokines are all necessary components. Co-stimulation regulates responsiveness of T cells and NKG2D is one of well-documented co-stimulatory molecules for T cells.,[40] CD28 –mediated co-stimulation is required for promotion of cytokine production and cytotoxicity in CD8+ T cells by NKG2D.[41] For cytokine production and cytolytic killing by γδ T cells, priming is not required, however NKG2D expression is constitutive while solely triggering NKG2D in γδ T cells does not mediate cytotoxicity.[42]
In T cells, NKG2D is associated with IL15-receptor signaling and also with the development of memory CD8 T cells.[43] Key role in the transformation of CD8+ T cells into effector or memory types is played by mammalian target of rapamycin complex 1 (mTORC1) – memory precursor cells are characteristic by the low level of mTORC1 and for terminally differentiated CD8+ T cells, high level of mTORC1 activity is typical.[44] Upregulation of antiapoptic Mcl-1 protein by NKG2D also induces formation of memory T cells.[43] In murine memory T cell precursors, downregulation of T-bet transcription factor is likewise affected by NKG2D.[45]
NKG2D and B cells
Also development of B cells is regulated by NKG2D—NKG2D deficient mice have reduced number of B cells in spleen,[46] which is partially depended on DAP10.[47] In comparison to NK cells, mature B cells do not express NKG2D.[46]
See also
References
- GRCh38: Ensembl release 89: ENSG00000213809 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Houchins JP, Yabe T, McSherry C, Bach FH (April 1991). "DNA sequence analysis of NKG2, a family of related cDNA clones encoding type II integral membrane proteins on human natural killer cells". The Journal of Experimental Medicine. 173 (4): 1017–20. doi:10.1084/jem.173.4.1017. PMC 2190798. PMID 2007850.
- Brown MG, Fulmek S, Matsumoto K, Cho R, Lyons PA, Levy ER, et al. (May 1997). "A 2-Mb YAC contig and physical map of the natural killer gene complex on mouse chromosome 6". Genomics. 42 (1): 16–25. doi:10.1006/geno.1997.4721. PMID 9177771.
- Yabe T, McSherry C, Bach FH, Fisch P, Schall RP, Sondel PM, Houchins JP (1993). "A multigene family on human chromosome 12 encodes natural killer-cell lectins". Immunogenetics. 37 (6): 455–60. doi:10.1007/BF00222470. PMID 8436421. S2CID 27350036.
- Jamieson AM, Diefenbach A, McMahon CW, Xiong N, Carlyle JR, Raulet DH (July 2002). "The role of the NKG2D immunoreceptor in immune cell activation and natural killing". Immunity. 17 (1): 19–29. doi:10.1016/S1074-7613(02)00333-3. PMID 12150888.
- Bauer S, Groh V, Wu J, Steinle A, Phillips JH, Lanier LL, Spies T (July 1999). "Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA". Science. 285 (5428): 727–9. doi:10.1126/science.285.5428.727. PMID 10426993.
- Raulet DH (October 2003). "Roles of the NKG2D immunoreceptor and its ligands". Nature Reviews. Immunology. 3 (10): 781–90. doi:10.1038/nri1199. PMID 14523385. S2CID 18234848.
- Li P, Morris DL, Willcox BE, Steinle A, Spies T, Strong RK (May 2001). "Complex structure of the activating immunoreceptor NKG2D and its MHC class I-like ligand MICA". Nature Immunology. 2 (5): 443–51. doi:10.1038/87757. PMID 11323699. S2CID 11096566.
- Garrity D, Call ME, Feng J, Wucherpfennig KW (May 2005). "The activating NKG2D receptor assembles in the membrane with two signaling dimers into a hexameric structure". Proceedings of the National Academy of Sciences of the United States of America. 102 (21): 7641–6. Bibcode:2005PNAS..102.7641G. doi:10.1073/pnas.0502439102. PMC 1140444. PMID 15894612.
- Upshaw JL, Arneson LN, Schoon RA, Dick CJ, Billadeau DD, Leibson PJ (May 2006). "NKG2D-mediated signaling requires a DAP10-bound Grb2-Vav1 intermediate and phosphatidylinositol-3-kinase in human natural killer cells". Nature Immunology. 7 (5): 524–32. doi:10.1038/ni1325. PMID 16582911. S2CID 28236529.
- Diefenbach A, Tomasello E, Lucas M, Jamieson AM, Hsia JK, Vivier E, Raulet DH (December 2002). "Selective associations with signaling proteins determine stimulatory versus costimulatory activity of NKG2D". Nature Immunology. 3 (12): 1142–9. doi:10.1038/ni858. PMID 12426565. S2CID 14901451.
- Gilfillan S, Ho EL, Cella M, Yokoyama WM, Colonna M (December 2002). "NKG2D recruits two distinct adapters to trigger NK cell activation and costimulation". Nature Immunology. 3 (12): 1150–5. doi:10.1038/ni857. PMID 12426564. S2CID 5859797.
- Raulet DH, Gasser S, Gowen BG, Deng W, Jung H (2013-01-01). "Regulation of ligands for the NKG2D activating receptor". Annual Review of Immunology. 31 (1): 413–41. doi:10.1146/annurev-immunol-032712-095951. PMC 4244079. PMID 23298206.
- Gasser S, Orsulic S, Brown EJ, Raulet DH (August 2005). "The DNA damage pathway regulates innate immune system ligands of the NKG2D receptor". Nature. 436 (7054): 1186–90. Bibcode:2005Natur.436.1186G. doi:10.1038/nature03884. PMC 1352168. PMID 15995699.
- Carapito R, Bahram S (September 2015). "Genetics, genomics, and evolutionary biology of NKG2D ligands". Immunological Reviews. 267 (1): 88–116. doi:10.1111/imr.12328. PMID 26284473. S2CID 205213312.
- Song P, Zhao Q, Zou MH (July 2020). "Targeting senescent cells to attenuate cardiovascular disease progression". Ageing Research Reviews. 60: 101072. doi:10.1016/j.arr.2020.101072. PMC 7263313. PMID 32298812.
- Cerboni C, Fionda C, Soriani A, Zingoni A, Doria M, Cippitelli M, Santoni A (January 2014). "The DNA Damage Response: A Common Pathway in the Regulation of NKG2D and DNAM-1 Ligand Expression in Normal, Infected, and Cancer Cells". Frontiers in Immunology. 4: 508. doi:10.3389/fimmu.2013.00508. PMC 3882864. PMID 24432022.
- Sancar A, Lindsey-Boltz LA, Unsal-Kaçmaz K, Linn S (June 2004). "Molecular mechanisms of mammalian DNA repair and the DNA damage checkpoints". Annual Review of Biochemistry. 73 (1): 39–85. doi:10.1146/annurev.biochem.73.011303.073723. PMID 15189136.
- Gavlovsky PJ, Tonnerre P, Gérard N, Nedellec S, Daman AW, McFarland BJ, Charreau B (August 2016). "Alternative Splice Transcripts for MHC Class I-like MICA Encode Novel NKG2D Ligands with Agonist or Antagonist Functions". Journal of Immunology. 197 (3): 736–46. doi:10.4049/jimmunol.1501416. PMID 27342847.
- Cao W, Xi X, Wang Z, Dong L, Hao Z, Cui L, et al. (August 2008). "Four novel ULBP splice variants are ligands for human NKG2D". International Immunology. 20 (8): 981–91. doi:10.1093/intimm/dxn057. PMID 18544572.
- Eagle RA, Flack G, Warford A, Martínez-Borra J, Jafferji I, Traherne JA, et al. (2009-02-18). "Cellular expression, trafficking, and function of two isoforms of human ULBP5/RAET1G". PLOS ONE. 4 (2): e4503. Bibcode:2009PLoSO...4.4503E. doi:10.1371/journal.pone.0004503. PMC 2637608. PMID 19223974.
- Vantourout P, Willcox C, Turner A, Swanson CM, Haque Y, Sobolev O, et al. (April 2014). "Immunological visibility: posttranscriptional regulation of human NKG2D ligands by the EGF receptor pathway". Science Translational Medicine. 6 (231): 231ra49. doi:10.1126/scitranslmed.3007579. PMC 3998197. PMID 24718859.
- Zingoni A, Molfetta R, Fionda C, Soriani A, Paolini R, Cippitelli M, et al. (2018-03-12). "NKG2D and Its Ligands: "One for All, All for One"". Frontiers in Immunology. 9: 476. doi:10.3389/fimmu.2018.00476. PMC 5890157. PMID 29662484.
- González S, López-Soto A, Suarez-Alvarez B, López-Vázquez A, López-Larrea C (August 2008). "NKG2D ligands: key targets of the immune response". Trends in Immunology. 29 (8): 397–403. doi:10.1016/j.it.2008.04.007. PMID 18602338.
- Zafirova B, Wensveen FM, Gulin M, Polić B (November 2011). "Regulation of immune cell function and differentiation by the NKG2D receptor". Cellular and Molecular Life Sciences. 68 (21): 3519–29. doi:10.1007/s00018-011-0797-0. PMC 3192283. PMID 21898152.
- Welte SA, Sinzger C, Lutz SZ, Singh-Jasuja H, Sampaio KL, Eknigk U, et al. (January 2003). "Selective intracellular retention of virally induced NKG2D ligands by the human cytomegalovirus UL16 glycoprotein". European Journal of Immunology. 33 (1): 194–203. doi:10.1002/immu.200390022. PMID 12594848. S2CID 20718868.
- Liu H, Wang S, Xin J, Wang J, Yao C, Zhang Z (2019). "Role of NKG2D and its ligands in cancer immunotherapy". American Journal of Cancer Research. 9 (10): 2064–2078. PMC 6834480. PMID 31720075.
- Serrano AE, Menares-Castillo E, Garrido-Tapia M, Ribeiro CH, Hernández CJ, Mendoza-Naranjo A, et al. (March 2011). "Interleukin 10 decreases MICA expression on melanoma cell surface". Immunology and Cell Biology. 89 (3): 447–57. doi:10.1038/icb.2010.100. hdl:10533/132162. PMID 20714339. S2CID 205150174.
- Sagiv A, Burton DG, Moshayev Z, Vadai E, Wensveen F, Ben-Dor S, et al. (February 2016). "NKG2D ligands mediate immunosurveillance of senescent cells". Aging. 8 (2): 328–44. doi:10.18632/aging.100897. PMC 4789586. PMID 26878797.
- Sagiv A, Biran A, Yon M, Simon J, Lowe SW, Krizhanovsky V. (2013). Granule exocytosis mediates immune surveillance of senescent cells Oncogene, 32, 1971–197, doi:10.1038/onc.2012.206
- Muñoz DP, Yannone SM, Daemen A, Sun Y, Vakar-Lopez F, Kawahara M, et al. (June 2019). "Targetable mechanisms driving immunoevasion of persistent senescent cells link chemotherapy-resistant cancer to aging". JCI Insight. 5 (14): 124716. doi:10.1172/jci.insight.124716. PMC 6675550. PMID 31184599.
- Dhar P, Wu JD (April 2018). "NKG2D and its ligands in cancer". Current Opinion in Immunology. 51: 55–61. doi:10.1016/j.coi.2018.02.004. PMC 6145810. PMID 29525346.
- Huntington ND, Vosshenrich CA, Di Santo JP (September 2007). "Developmental pathways that generate natural-killer-cell diversity in mice and humans". Nature Reviews. Immunology. 7 (9): 703–14. doi:10.1038/nri2154. PMID 17717540.
- Rabinovich B, Li J, Wolfson M, Lawrence W, Beers C, Chalupny J, et al. (April 2006). "NKG2D splice variants: a reexamination of adaptor molecule associations". Immunogenetics. 58 (2–3): 81–8. doi:10.1007/s00251-005-0078-x. PMID 16470377. S2CID 20895192.
- Di Santo JP (April 2006). "Natural killer cell developmental pathways: a question of balance". Annual Review of Immunology. 24 (1): 257–86. doi:10.1146/annurev.immunol.24.021605.090700. PMID 16551250.
- Horng T, Bezbradica JS, Medzhitov R (December 2007). "NKG2D signaling is coupled to the interleukin 15 receptor signaling pathway". Nature Immunology. 8 (12): 1345–52. doi:10.1038/ni1524. PMID 17952078. S2CID 13356420.
- Groh V, Rhinehart R, Secrist H, Bauer S, Grabstein KH, Spies T (June 1999). "Broad tumor-associated expression and recognition by tumor-derived gamma delta T cells of MICA and MICB". Proceedings of the National Academy of Sciences of the United States of America. 96 (12): 6879–84. Bibcode:1999PNAS...96.6879G. doi:10.1073/pnas.96.12.6879. PMC 22010. PMID 10359807.
- Hedlund M, Stenqvist AC, Nagaeva O, Kjellberg L, Wulff M, Baranov V, Mincheva-Nilsson L (July 2009). "Human placenta expresses and secretes NKG2D ligands via exosomes that down-modulate the cognate receptor expression: evidence for immunosuppressive function". Journal of Immunology. 183 (1): 340–51. doi:10.4049/jimmunol.0803477. PMID 19542445.
- Groh V, Rhinehart R, Randolph-Habecker J, Topp MS, Riddell SR, Spies T (March 2001). "Costimulation of CD8alphabeta T cells by NKG2D via engagement by MIC induced on virus-infected cells". Nature Immunology. 2 (3): 255–60. doi:10.1038/85321. PMID 11224526. S2CID 35460567.
- Kavazović I, Lenartić M, Jelenčić V, Jurković S, Lemmermann NA, Jonjić S, et al. (July 2017). "NKG2D stimulation of CD8+ T cells during priming promotes their capacity to produce cytokines in response to viral infection in mice". European Journal of Immunology. 47 (7): 1123–1135. doi:10.1002/eji.201646805. PMID 28378389.
- Nedellec S, Sabourin C, Bonneville M, Scotet E (July 2010). "NKG2D costimulates human V gamma 9V delta 2 T cell antitumor cytotoxicity through protein kinase C theta-dependent modulation of early TCR-induced calcium and transduction signals". Journal of Immunology. 185 (1): 55–63. doi:10.4049/jimmunol.1000373. PMID 20511557.
- Wensveen FM, Lenartic M, Jelencic V, Lemmermann NA, ten Brinke A, Jonjic S, Polic B (August 2013). "NKG2D induces Mcl-1 expression and mediates survival of CD8 memory T cell precursors via phosphatidylinositol 3-kinase". Journal of Immunology. 191 (3): 1307–15. doi:10.4049/jimmunol.1300670. PMID 23804716.
- McQueen B, Trace K, Whitman E, Bedsworth T, Barber A (March 2016). "Natural killer group 2D and CD28 receptors differentially activate mammalian/mechanistic target of rapamycin to alter murine effector CD8+ T-cell differentiation". Immunology. 147 (3): 305–20. doi:10.1111/imm.12563. PMC 4754608. PMID 26661515.
- Zloza A, Kohlhapp FJ, Lyons GE, Schenkel JM, Moore TV, Lacek AT, et al. (February 2012). "NKG2D signaling on CD8⁺ T cells represses T-bet and rescues CD4-unhelped CD8⁺ T cell memory recall but not effector responses". Nature Medicine. 18 (3): 422–8. doi:10.1038/nm.2683. PMC 3436127. PMID 22366950.
- Zafirova B, Mandarić S, Antulov R, Krmpotić A, Jonsson H, Yokoyama WM, et al. (August 2009). "Altered NK cell development and enhanced NK cell-mediated resistance to mouse cytomegalovirus in NKG2D-deficient mice". Immunity. 31 (2): 270–82. doi:10.1016/j.immuni.2009.06.017. PMC 2782462. PMID 19631564.
- Wensveen FM, Jelenčić V, Polić B (2018-03-08). "NKG2D: A Master Regulator of Immune Cell Responsiveness". Frontiers in Immunology. 9: 441. doi:10.3389/fimmu.2018.00441. PMC 5852076. PMID 29568297.