Haplogroup A (mtDNA)
In human mitochondrial genetics, Haplogroup A is a human mitochondrial DNA (mtDNA) haplogroup.
| Haplogroup A | |
|---|---|
 | |
| Possible time of origin | 40,000 ± 10,000 YBP 40,500 (95% CI 37,900 <-> 43,200) ybp[1] |
| Coalescence age | 18,600 (95% CI 14,200 <-> 23,900) ybp[1] 24,209 (SD 4,906) ybp[2] |
| Possible place of origin | Asia |
| Ancestor | N |
| Descendants | A3, A4, A5, A7, A8 |
| Defining mutations | 152, 235, 523-524d, 663, 1736, 4248, 4824, 8794, 16290, 16319[3] |
Origin
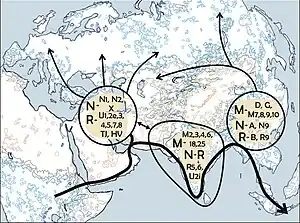
Haplogroup A is believed to have arisen in Asia some 30,000–50,000 years BC. Its ancestral haplogroup was Haplogroup N. However, the extant diversity of mitochondrial genomes that belong to Haplogroup A is low relative to the degree of divergence from its nearest outgroups in haplogroup N, which suggests that extant members of Haplogroup A might be descended from a population that has emerged from a bottleneck approximately 20,000 years ago.
Its highest frequencies are among Native Americans, its largest overall population is in East Asia, and its greatest variety (which suggests its origin point) is in East Asia. Thus, it might have originated in and spread from the Far East.[4]
Distribution
Its subclade A2 shares a T16362C mutation with subclades A1 (found in Japan, Tashkurgan, Veliky Novgorod, Mongols, and Altaians), A6 (found in Tibet and in the Yangtze River basin), A12'23 (found in Siberia and among Uralic and Turkic peoples), A13'14 (found in southern Siberia, Xinjiang, Ladakh, China, Yunnan, Thailand, and Vietnam), A15 (found in China, Naxi, Uyghur, Japan, and among the Sherpa of Tibet and Nepal), A16 (found in Uyghur, Buryat, Turkey), A17 (found in China, Miao, Yi, Tibet, Ladakh, Kyrgyz, Thailand, and Vietnam), A18 (found in China), A19 (found in China), A20 (found among Han Chinese and in Japan), A21 (found in Tibet and in Jammu and Kashmir), A22 (found in China), A24 (found in Beijing and West Bohemia), A25 (found in Japan and Yakutia), and A26 (found in Denmark). A2 is found in Chukotko–Kamchatka[5] and is also one of five mtDNA haplogroups found in the indigenous peoples of the Americas, the others being B, C, D, and X.[4]
Haplogroup A2 is the most common haplogroup among the Inuit, Na-Dene, and many Amerind ethnic groups of North and Central America. Lineages belonging to haplogroup A2 also comprise the majority of the mtDNA pool of the Inuit and their neighbors, the Chukchis, in northeasternmost Siberia.[5][6][7]
Other branches of haplogroup A are less frequent but widespread among other populations of Asia.[8][9] Haplogroup A5 is rather limited to populations from Korea and Japan southward, though it has been detected as singletons in a pair of large samples of Khamnigans (1/99 = 1.0%) and Buryats (1/295 = 0.3%) from the Buryat Republic.[6]
In Asia, A(xA2) is especially frequent in Tibeto-Burman-speaking populations of Southwest China, such as Tibetans (6/65 = 9.2%,[5] 25/216 = 11.6%,[10] 11/73 = 15.1%[10]). Approximately 7% to 15% of Koreans belong to haplogroup A.[6][11][12] Approximately 5% to 12% of the Japanese belong to haplogroup A (including A4, A5, and A(xA4, A5)).[5][13][14][15] Approximately 4% to 13% of Mongols belong to haplogroup A, almost all of whom are contained within the A4 subclade (2/47 = 4.3% Mongolians from Ulan Bator in haplogroup A4,[11] 4/48 = 8.3% Mongols from New Barag Left Banner in haplogroup A(xA5),[12] 6/47 = 12.8% Mongolians from Ulan Bator in haplogroup A4[6]). Approximately 3% to 9% of Chinese people belong to haplogroup A.[13] Haplogroup A also has been found in Vietnamese (2/42 = 4.8%, including one A4 and one A5(xA5a)).[11] Approximately 4% (3/71) of Tatars from Aznakayevo,[16] 3% (4/126) of Tatars from Buinsk,[16] and 2% of Turkish people belong to haplogroup A.[17] Haplogroup A4 has been found in 2.4% (2/82) of a sample of Persians from eastern Iran and in 2.3% (1/44) of a sample of Tajiks from Tajikistan.[6] Haplogroup A is not found among Austronesians.[18] In Nepalese population except Sherpa, haplogroup A was mirrored by its clades, A27, A14 and A17, of which A27 was the most abundant clade in Newar (3.99%).[19] Newly defined clade A27 only discerned so far in Newar and Nepali-mix coalesce at ~ 8.4 Kya suggesting their ancient origin and potentially insitu differentiation in Nepal.[19]
Table of Frequencies of MtDNA Haplogroup A
| Population | Frequency | Count | Source | Subtypes |
|---|---|---|---|---|
| Tłı̨chǫ (Dogrib) | 1 | 42 | [20] | |
| Tlingit | 1 | 2 | [20] | |
| Acoma Pueblo | 1 | 1 | [20] | |
| Esselen | 1 | 1 | [21] | A01 |
| Haida | 0.966 | 29 | [20] | |
| Eskimo (Greenland) | 0.961 | 385 | Volodko 2008 | A2b=196, A2a=174 |
| Eskimo (Chaplin) | 0.900 | 50 | Volodko 2008 | A2a=36, A2b=9 |
| Eskimo (Canada) | 0.875 | 96 | Volodko 2008 | A2b=68, A2a=16 |
| Mixtec | 0.828 | 29 | [20] | |
| Siberian Eskimo | 0.772 | 79 | A2=61 (41/46 Chaplin, 17/25 Sireniki, 3/8 Naukan) | |
| Eskimo (Naukan) | 0.744 | 39 | Volodko 2008 | A2b=16, A2a=13 |
| Chukchi (Anadyr, Chukotka) | 0.733 | 15 | [6] | A2=11 |
| Eskimo (Sireniki) | 0.703 | 37 | Volodko 2008 | A2a=16, A2b=10 |
| Chukchi | 0.682 | 66 | A2=45 | |
| Chickasaw/Choctaw | 0.667 | 27 | [20] | |
| Mixe | 0.625 | 16 | [20] | |
| Apache | 0.621 | 29 | [20] | |
| Nahua (Cuetzalan, Mexico) | 0.613 | 31 | A=19 | |
| Nahua/Cora (Mexico) | 0.531 | 32 | [20] | |
| Siouan | 0.529 | 34 | [20] | |
| Chumash | 0.524 | 21 | [21] | A02, A03, A04, A05, A07, A09, A10, A12 |
| Maya (Mexico) | 0.519 | 27 | [20] | |
| Navajo | 0.517 | 58 | [20] | |
| Nuxalk (Bella Coola) | 0.5 | 36 | [20] | |
| Salinan | 0.5 | 6 | [21] | A01, A06, A13 |
| Ojibwe (Chippewa)/Kickapoo | 0.484 | 62 | [20] | |
| Salinan/Chumash | 0.455 | 11 | [20] | |
| Nuu-Chah-Nulth | 0.4 | 15 | [20] | |
| Kiowa | 0.4 | 5 | [20] | |
| Creek/Seminole | 0.389 | 18 | [20] | |
| Aleut (Aleutian Islands) | 0.344 | 163 | Volodko 2008 | A2a=56 |
| Zapotec | 0.333 | 15 | [20] | |
| Pawnee | 0.333 | 3 | [20] | |
| Cheyenne/Arapaho | 0.308 | 26 | [20] | |
| Nu (Gongshan, Yunnan) | 0.300 | 30 | A=9 | |
| Lisu (Gongshan, Yunnan) | 0.297 | 37 | A=11 | |
| Mi'kmaq (Newfoundland)/Narragansett | 0.286 | 7 | [20] | |
| Chuvantsi (Markovo, Chukotka) | 0.250 | 32 | Volodko 2008 | A2a=6, A2b=2 |
| Tibetan (Diqing, Yunnan) | 0.250 | 24 | A=6 | |
| Yi (Hezhang County, Guizhou) | 0.250 | 20 | A=5 | |
| Ohlone (Costanoan) | 0.25 | 8 | [23] | A01 |
| Tibetan (Nagchu, Tibet) | 0.229 | 35 | A=8 | |
| Tibetan (Qinghai) | 0.214 | 56 | A=12 | |
| Tibetan (Shannan, Tibet) | 0.211 | 19 | A=4 | |
| Yi (Xishuangbanna, Yunnan) | 0.188 | 16 | A=3 | |
| Tibetan (Chamdo, Tibet) | 0.172 | 29 | A1=5 | |
| Zuni | 0.182 | 22 | [20] | |
| Korean (Arun Banner) | 0.146 | 48 | [12] | A5=4, A(xA5)=3 |
| Tujia (Western Hunan) | 0.141 | 64 | A=9 | |
| Pumi (Ninglang, Yunnan) | 0.139 | 36 | A=5 | |
| Tujia (Yanhe County, Guizhou) | 0.138 | 29 | A=4 | |
| Tibetan (Lhasa, Tibet) | 0.136 | 44 | A1=6 | |
| Mongolian (Ulan Bator) | 0.128 | 47 | [6] | A4(xA2)=6 |
| Hani (Xishuangbanna, Yunnan) | 0.121 | 33 | A=4 | |
| Japanese (Miyazaki) | 0.120 | 100 | A4=4, A5=4, A(xA4,A5)=4 | |
| Gelao (Daozhen County, Guizhou) | 0.118 | 102 | A=12 | |
| Penutian (California) | 0.118 | 17 | [20] | |
| Tibetan (Zhongdian, Yunnan) | 0.114 | 35 | A=4 | |
| Tubalar (Turochak & Choysky) | 0.111 | 72 | A(xA2)=8 | |
| Havasupai/Hualapai/Yavapai/Mojave | 0.111 | 18 | [20] | |
| Tibetan (Shannan, Tibet) | 0.109 | 55 | A1=6 | |
| Tibetan (Shigatse, Tibet) | 0.103 | 29 | A1=3 | |
| Yi (Shuangbai, Yunnan) | 0.100 | 40 | A=4 | |
| Manchurian | 0.100 | 40 | [11] | A(xA4,A5)=3, A4=1 |
| Korean (northern China) | 0.098 | 51 | [11] | A4=4, A5(xA5a)=1 |
| Yi (Luxi, Yunnan) | 0.097 | 31 | A=3 | |
| Han (Denver) | 0.096 | 73 | Zheng 2011 | A=7 |
| Japanese | 0.090 | 211 | A5=11, A(xA5)=8 | |
| Naxi (Lijiang, Yunnan) | 0.089 | 45 | A=4 | |
| Korean (South Korea) | 0.089 | 203 | [13] | A=18 |
| Chinese (Shenyang, Liaoning) | 0.088 | 160 | [13] | A=14 |
| Hmong (Jishou, Hunan) | 0.087 | 103 | A(xA6)=7, A6=2 | |
| Japanese (Tōhoku) | 0.086 | 336 | [13] | A=29 |
| Mongol (New Barag Left Banner) | 0.083 | 48 | [12] | A(xA5)=4 |
| Korean (South Korea) | 0.081 | 185 | [11] | A4=6, A5(xA5a)=5, A(xA4,A5)=3, A5a=1 |
| Cochimí | 0.077 | 13 | [20] | |
| Korean (South Korea) | 0.077 | 261 | A=20 | |
| Han (Beijing Normal University) | 0.074 | 121 | Zheng 2011 | A=9 |
| Pai Yuman | 0.074 | 27 | A=2 | |
| Tibetan (Nyingchi, Tibet) | 0.074 | 54 | A1=4 | |
| Han (Southwest China, pool of 44 Sichuan, 34 Chongqing, 33 Yunnan, and 26 Guizhou) | 0.073 | 137 | A=10 | |
| Han (Hunan and Fujian) | 0.073 | 55 | Zheng 2011 | A=4 |
| Telengit | 0.073 | 55 | A=4 | |
| Korean (Seoul National University Hospital) | 0.073 | 633 | Fuku 2007 | A=46 |
| Buryat | 0.071 | 126 | [12] | A(xA5)=9 |
| Han (southern California) | 0.069 | 390 | A=27 | |
| Korean (South Korea) | 0.068 | 103 | [6] | A5=4, A4(xA2)=3 |
| Japanese (Tokyo) | 0.068 | 118 | Zheng 2011 | A=8 |
| Okinawa | 0.067 | 326 | [13] | A=22 |
| Japanese (northern Kyūshū) | 0.066 | 256 | [13] | A=17 |
| Itelmen | 0.064 | 47 | A(xA2)=3 | |
| Japanese (Gifu) | 0.063 | 1617 | Fuku 2007 | A=102 |
| Yokuts | 0.063 | 16 | [21] | A08 |
| Zhuang (Napo County, Guangxi) | 0.062 | 130 | A=8 | |
| Barghut (Hulun Buir) | 0.060 | 149 | A4=8, A8=1 | |
| Japanese (Hokkaidō) | 0.060 | 217 | Asari 2007 | A=13 |
| Bai (Dali, Yunnan) | 0.059 | 68 | A=4 | |
| Ket | 0.059 | 34 | [24] | A8a2 |
| Evenk (Siberia) | 0.056 | 71 | A(xA2)=4 | |
| Telenghit (Altai Republic) | 0.056 | 71 | [6] | A4(xA2)=4 |
| Jino (Xishuangbanna, Yunnan) | 0.056 | 18 | A=1 | |
| Bai (Xishuangbanna, Yunnan) | 0.053 | 19 | A=1 | |
| Koryak | 0.052 | 155 | A2=4, A(xA2)=4 | |
| Buryat (Buryatia) | 0.051 | 295 | [6] | A4(xA2)=13, A5=1, A8=1 |
| Khamnigan (Buryatia) | 0.051 | 99 | [6] | A4(xA2)=4, A5=1 |
| Tibetan (Deqin, Yunnan) | 0.050 | 40 | A=2 | |
| Han (Beijing) | 0.050 | 40 | [11] | A4=1, A(xA4,A5)=1 |
| Japanese (Tōkai) | 0.050 | 282 | [13] | A=14 |
| Dai (Xishuangbanna, Yunnan) | 0.049 | 41 | A=2 | |
| Vietnamese | 0.048 | 42 | [11] | A4=1, A5(xA5a)=1 |
| Yakama | 0.048 | 42 | A=2 | |
| Akimal O’odham (Pima) | 0.054 | 37 | [20] | |
| Han (Kunming, Yunnan) | 0.047 | 43 | A=2 | |
| Dolgan (Anabarsky, Volochanka, Ust-Avam, & Dudinka) | 0.045 | 154 | A10=3, A8=2, A4(xA4b)=2 | |
| Oroqen (Oroqen Autonomous Banner) | 0.045 | 44 | [12] | A(xA5)=2 |
| Va (Simao, Yunnan) | 0.045 | 22 | A=1 | |
| Evenk (New Barag Left Banner) | 0.043 | 47 | [12] | A(xA5)=2 |
| Mongolian (Ulan Bator) | 0.043 | 47 | [11] | A4=2 |
| Tatar (Aznakayevo) | 0.042 | 71 | Malyarchuk 2010 | A(xA8b)=2, A8b=1 |
| Altai-kizhi | 0.042 | 48 | A=2 | |
| Guoshan Yao (Jianghua, Hunan) | 0.042 | 24 | A(xA6)=1 | |
| Evenk (Krasnoyarsk) | 0.041 | 73 | [6] | A4(xA2)=3 |
| Evenk (Ust-Maysky, Oleneksky, Zhigansky) | 0.040 | 125 | A4(xA4b)=3, A4b=2 | |
| Ainu | 0.039 | 51 | Sato 2009[25] | A=2 |
| Kalmyk (Kalmykia) | 0.036 | 110 | [6] | A4(xA2)=3, A8=1 |
| Han (Taiwanese) | 0.036 | 111 | A4e1=2, A5b=2 | |
| Yakut (Vilyuy River basin) | 0.036 | 111 | A4(xA4b)=2, A4b=1, A8=1 | |
| Han (Taiwan) | 0.036 | 1117 | A=40 | |
| Dong (Tianzhu County, Guizhou) | 0.036 | 28 | A=1 | |
| Shor | 0.036 | 28 | A=1 | |
| Khakassian (Khakassia) | 0.035 | 57 | [6] | A4(xA2)=2 |
| Altay Kizhi | 0.033 | 90 | [6] | A4(xA2)=3 |
| Taiwanese (Taipei, Taiwan) | 0.033 | 91 | [13] | A=3 |
| Wuzhou Yao (Fuchuan, Guangxi) | 0.032 | 31 | A(xA6)=1 | |
| Tatar (Buinsk) | 0.032 | 126 | Malyarchuk 2010 | A8b=4 |
| Pan Yao (Tianlin, Guangxi) | 0.031 | 32 | A6=1 | |
| Kazakh (Kosh-Agach District) | 0.031 | 98 | A4=3 | |
| Mansi | 0.031 | 98 | A(xA2)=3 | |
| Altai-kizhi (Altai Republic) | 0.029 | 276 | A=8 | |
| Bapai Yao (Liannan, Guangdong) | 0.029 | 35 | A6=1 | |
| Guangdong | 0.026 | 546 | A=14 | |
| Kim Mun (Malipo, Yunnan) | 0.025 | 40 | A6=1 | |
| Persian (eastern Iran) | 0.024 | 82 | [6] | A4(xA2)=2 |
| Tu Yao (Hezhou, Guangxi) | 0.024 | 41 | A6=1 | |
| Yakut (vicinity of Yakutsk) | 0.024 | 164 | A4b=2, A4(xA4b)=1, A8=1 | |
| Lowland Yao (Fuchuan, Guangxi) | 0.024 | 42 | A(xA6)=1 | |
| Tajik (Tajikistan) | 0.023 | 44 | [6] | A4(xA2)=1 |
| Daur (Evenk Autonomous Banner) | 0.022 | 45 | [12] | A(xA5)=1 |
| Evenk (Buryatia) | 0.022 | 45 | [6] | A4(xA2)=1 |
| Tuvan | 0.021 | 95 | A(xA2)=2 | |
| Aini (Xishuangbanna, Yunnan) | 0.020 | 50 | A=1 | |
| Kumandin (Turochak District) | 0.019 | 52 | A=1 | |
| Guangxi | 0.017 | 1111 | A=19 | |
| Yakut | 0.017 | 117 | [12] | A(xA5)=2 |
| Shor (Kemerovo) | 0.012 | 82 | [6] | A4(xA2)=1 |
| Tuvinian (Tuva) | 0.010 | 105 | [6] | A4(xA2)=1 |
| Khanty | 0.009 | 106 | [8] | A=1 |
| Vietnam | 0.008 | 392 | A=3 | |
| Southeast Yunnan | 0.006 | 158 | A=1 | |
| Li (Hainan) | 0.003 | 346 | A=1 | |
| Kiliwa | 0.000 | 7 | – | |
| Seri | 0.000 | 8 | – | |
| Paiute/Shoshone | 0 | 9 | [20] | – |
| Dingban Yao (Mengla, Yunnan) | 0.000 | 10 | – | |
| Xiban Yao (Fangcheng, Guangxi) | 0.000 | 11 | – | |
| Kiliwa/Paipai | 0 | 11 | [20] | – |
| Uto-Aztecan (California) | 0 | 14 | [20] | |
| Lahu (Xishuangbanna, Yunnan) | 0.000 | 15 | – | |
| Kumeyaay | 0 | 16 | [20] | – |
| Yukaghir (Upper Kolyma) | 0.000 | 18 | Volodko 2008 | – |
| Huatou Yao (Fangcheng, Guangxi) | 0.000 | 19 | – | |
| Filipino (Palawan) | 0.000 | 20 | – | |
| Dai (Xishuangbanna, Yunnan) | 0.000 | 21 | – | |
| Yukaghir (Verkhnekolymsky & Nizhnekolymsky) | 0.000 | 22 | – | |
| River Yuman | 0.000 | 22 | – | |
| Delta Yuman | 0.000 | 23 | – | |
| Quechan/Cocopah | 0 | 23 | [20] | – |
| Hindu (Chitwan, Nepal) | 0.000 | 24 | – | |
| Nganasan | 0.000 | 24 | – | |
| Tibetan (Nyingchi, Tibet) | 0.000 | 24 | – | |
| Buryat (Kushun, Nizhneudinsk, Irkutsk) | 0.000 | 25 | – | |
| Bunu (Dahua & Tianlin, Guangxi) | 0.000 | 25 | – | |
| Kurd (northwestern Iran) | 0.000 | 25 | [6] | – |
| Lanten Yao (Tianlin, Guangxi) | 0.000 | 26 | – | |
| Iu Mien (Mengla, Yunnan) | 0.000 | 27 | – | |
| Washo | 0 | 28 | [20] | – |
| Andhra Pradesh (tribal) | 0.000 | 29 | – | |
| Batek (Malaysia) | 0.000 | 29 | – | |
| Cun (Hainan) | 0.000 | 30 | – | |
| Tujia (Yongshun, Hunan) | 0.000 | 30 | – | |
| Batak (Palawan) | 0.000 | 31 | – | |
| Gelao (Daozhen County, Guizhou) | 0.000 | 31 | – | |
| Lingao (Hainan) | 0.000 | 31 | – | |
| Lahu (Simao, Yunnan) | 0.000 | 32 | – | |
| Mendriq (Malaysia) | 0.000 | 32 | – | |
| Mien (Shangsi, Guangxi) | 0.000 | 32 | – | |
| Negidal | 0.000 | 33 | – | |
| Teleut | 0.000 | 33 | – | |
| Temuan (Malaysia) | 0.000 | 33 | – | |
| Lahu (Lancang, Yunnan) | 0.000 | 35 | – | |
| Aleut (Commander Islands) | 0.000 | 36 | Volodko 2008 | – |
| Va (Ximeng & Gengma, Yunnan) | 0.000 | 36 | – | |
| Yakut (Yakutia) | 0.000 | 36 | [6] | – |
| Jemez/Taos/San Ildefonso Pueblo | 0 | 36 | [20] | – |
| Taono O’odham | 0.000 | 37 | – | |
| Hmong (Wenshan, Yunnan) | 0.000 | 39 | – | |
| Nganasan | 0.000 | 39 | Volodko 2008 | – |
| Thai | 0.000 | 40 | [11] | – |
| Tharu (Morang, Nepal) | 0.000 | 40 | – | |
| Ambon | 0.000 | 43 | – | |
| Lombok (Mataram) | 0.000 | 44 | – | |
| Alor | 0.000 | 45 | – | |
| Tofalar | 0.000 | 46 | – | |
| Udegey | 0.000 | 46 | – | |
| Hindu (New Delhi, India) | 0.000 | 48 | – | |
| Sumba (Waingapu) | 0.000 | 50 | – | |
| Jahai (Malaysia) | 0.000 | 51 | – | |
| Senoi (Malaysia) | 0.000 | 52 | – | |
| Teleut (Kemerovo) | 0.000 | 53 | [6] | – |
| Nivkh (northern Sakhalin) | 0.000 | 56 | – | |
| Filipino | 0.000 | 61 | – | |
| Semelai (Malaysia) | 0.000 | 61 | – | |
| Mansi | 0.000 | 63 | [8] | – |
| Filipino | 0.000 | 64 | [18] | – |
| Filipino (Mindanao) | 0.000 | 70 | [18] | – |
| Tubalar (Turochak District) | 0.000 | 71 | – | |
| Bali | 0.000 | 82 | – | |
| Yukaghir (Lower Kolyma-Indigirka) | 0.000 | 82 | Volodko 2008 | – |
| Ulchi | 0.000 | 87 | – | |
| Chelkan (Turochak District) | 0.000 | 91 | – | |
| N. Paiute/Shoshoni | 0.000 | 94 | – | |
| Northern Paiute | 0.000 | 98 | – | |
| Even (Eveno-Bytantaysky & Momsky) | 0.000 | 105 | – | |
| Tharu (Chitwan, Nepal) | 0.000 | 133 | – | |
| Yakut (northern Yakutia) | 0.000 | 148 | – | |
| Cham (Bình Thuận, Vietnam) | 0.000 | 168 | – | |
| Filipino (Luzon) | 0.000 | 177 | [18] | – |
| Sumatra | 0.000 | 180 | – | |
| Sulawesi | 0.000 | 237 | – | |
| Taiwan aborigine | 0.000 | 640 | – | |
Subclades
Tree
This phylogenetic tree of haplogroup A subclades is based on the paper by Mannis van Oven and Manfred Kayser Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation[3] and subsequent published research.
- A
- A(xA5, A8, A10) – China (Han from Wuhan), Buryat (Inner Mongolia)
- A+T152C!+T16362C – Uyghur, Korea, Japan, Vietnam (Hmong from Lao Cai Province,[26] Kinh from Hanoi,[26] Cờ Lao)
- A1 [TMRCA 12,800 (95% CI 6,500 <-> 22,700) ybp[1]]
- A1* – Japan, Korea
- A1a [TMRCA 7,500 (95% CI 4,500 <-> 11,800) ybp[1]]
- A1a* – Japan (Aichi), Sarikoli (Tashkurgan), USA, England
- A1a1 [TMRCA 5,000 (95% CI 2,200 <-> 9,800) ybp[1]]
- A1a1* – Buryat, Altai Kizhi
- A1a1a – Buryat, Mongol (Inner Mongolia) [TMRCA 1,050 (95% CI 75 <-> 5,500) ybp[1]]
- A1a2 – Russia (Bashkortostan, Velikij Novgorod), Iran (Turkmen) [TMRCA 1,950 (95% CI 100 <-> 10,500) ybp[1]]
- A1a3 – Greece (Ioannina), United States (West Virginia) [TMRCA 1,150 (95% CI 75 <-> 6,000) ybp[1]]
- A2 – Ache, Waiwai, Zoro, Surui, Waiapi, Poturujara, Kayapo, Katuena, Guarani, Arsario, Cayapa, Dogrib, ancient Canada, USA (Pennsylvania, California), Mexico (Zapotec), Cuba, Dominican Republic, Colombia, Venezuela, Ecuador, Peru, Argentina [TMRCA 10,600 (95% CI 9,600 <-> 11,700) ybp[1]]
- A2a – Eskimo (Greenland, Chukotka), Chukchi
- A2a1 – Inuit (Canada), Selkup[27]
- A2a2 – Eskimo (Chukotka), Chukchi
- A2a3 – Eskimo (Greenland, Canada, Chukotka), Chukchi
- A2a4 – USA (New Mexico, Arizona), Mexico (Chihuahua)
- A2a5 – Apache, USA (California, Arizona, New Mexico, Texas), Canada (Cree, Shuswap)
- A2b – Chukchi
- A2b1 – Chukchi, Koryak, Eskimo (Chukotka, Canada, Greenland)
- A2c
- A2d – USA (Mexican, Hispanic)
- A2d1 – USA (Mexican)
- A2d1a – USA (Hispanic)
- A2d2 – USA (Hispanic)
- A2d1 – USA (Mexican)
- A2e'ao
- A2e
- A2ao
- A2ao1
- A2f
- A2f1 – Newfoundland
- A2f1a – Canada, USA (Native American)
- A2f2 – USA (Mexican, Hispanic), Mexico
- A2f3 – USA (Mexican, Hispanic)
- A2f1 – Newfoundland
- A2g – USA (Mexican, Hispanic), Mexico, Iberian Peninsula
- A2g1 – USA (Mexican, Hispanic), Latin America
- A2h – Colombia (Cocama of Amazonas, Arhuaco), Yanomama, Kogui
- A2h1 – USA (Mexican, Hispanic), Mexico, Latin America
- A2i – USA (Hispanic, etc.), Canada (Ojibwa, Prince Edward Island, Pabos in Quebec)
- A2j – USA (Hispanic)
- A2j1 – USA (Hispanic)
- A2k – USA (Puerto Rico)
- A2k1 – Ecuador, Wayuu, Mexico
- A2k1a – Venezuela, Colombia (Pasto of Putumayo), USA (Hispanic)
- A2k1 – Ecuador, Wayuu, Mexico
- A2l'm'n'o'ai'aj
- A2l
- A2m
- A2n – Canada
- A2o
- A2ai
- A2aj
- A2p'am
- A2q
- A2q1
- A2r – USA (Hispanic, Mexican), Cuba
- A2r1 – Mexico, USA (Mexican)
- A2s
- A2t – USA (Mexican)
- A2u
- A2u1
- A2u2
- A2v
- A2v1 – USA (Mexican, Hispanic), Mexico (La Mixteca)
- A2v1a – Guatemala, USA (Mexican)
- A2v1b – Mexico
- A2v1 – USA (Mexican, Hispanic), Mexico (La Mixteca)
- A2w – Colombia (Kogi, Guambiano of Putumayo), Arsario, USA (Mexican, Hispanic)
- A2w1 – Mexico, Cayman Islands, Guatemala (La Tinta), Panama (Guaymi), Colombia
- A2x
- A2y
- A2z – USA (Hispanic, Puerto Rico)
- A2aa
- A2ab
- A2ac
- A2ac1
- A2ad
- A2ad1
- A2ad2
- A2ae
- A2af
- A2af1
- A2af1a
- A2af1a1
- A2af1a2
- A2af1b
- A2af1b1
- A2af1b1a
- A2af1b1b
- A2af1b2
- A2af1b1
- A2af1a
- A2af2
- A2af1
- A2ag
- A2ah
- A2ak
- A2al
- A2an
- A2ap
- A2aq
- A2a – Eskimo (Greenland, Chukotka), Chukchi
- A6 [TMRCA 12,000 (95% CI 8,600 <-> 16,100) ybp[1]]
- A12'23 – Austria, Romania, Poland, Russia, possibly found among Udmurts and Komis[27]
- A12 – Czech Republic, Germany [TMRCA 11,800 (95% CI 6,500 <-> 19,700) ybp[1]]
- A12a – Ireland, UK, New Zealand, USA, Nenets,[27] Selkup[27] [TMRCA 4,700 (95% CI 2,700 <-> 7,600) ybp[1]]
- A12a* – Mansi, Yakut (Vilyuy River basin),[30] Kyrgyz (Kyrgyzstan)[31]
- A12a1 – Kyordyughen Site (Ymyiakhtakh Culture, Yakutia),[32] Hungary (Debrecen) [TMRCA 2,800 (95% CI 1,450 <-> 4,900) ybp[1]]
- A12a2 – Evenk (Krasnoyarsk Krai,[6] Stony Tunguska River basin[30]) [TMRCA 1,250 (95% CI 100 <-> 6,600) ybp[1]]
- A12b – Buryat,[6] Karos-Eperjesszög (Hungarian conqueror period)[33] [TMRCA 3,000 (95% CI 425 <-> 10,700) ybp[1]]
- A12a – Ireland, UK, New Zealand, USA, Nenets,[27] Selkup[27] [TMRCA 4,700 (95% CI 2,700 <-> 7,600) ybp[1]]
- A23 – Mongol (Inner Mongolia),[34] Buryat,[6] Ket,[27] Qashqai (Iran),[35] ancient Scythian (Chylenski) [TMRCA 6,200 (95% CI 3,300 <-> 10,600) ybp[1]]
- A12 – Czech Republic, Germany [TMRCA 11,800 (95% CI 6,500 <-> 19,700) ybp[1]]
- A13'14 – Russia (Buryat, Khamnigan), China (Shiyan, Tu, Uyghur, etc.), Ladakh, Thailand, Vietnam (Mang), Korea, Japan, Paraguay (Alto Parana[36][1]), Ireland
- A13
- A13a - Thailand (Khon Mueang from Chiang Rai Province and Lampang Province[37][1]), China[1]
- A13b
- A14 – Russia (Altai Kizhi, etc.), Kyrgyz (Artux), Uyghur, China, Han Chinese (Denver), Taiwan, Thailand (Lawa from Chiang Mai Province, Mon from Lopburi Province[37]), Vietnam (Pa Then)
- A13
- A15 – Uyghur
- A16 – Buryat, Uyghur, Turk
- A17 – China (Han from Beijing, Lanzhou,[39] etc.), Miao, Yi, Tibet (Lhoba, Monpa, Tingri), Ladakh, Kyrgyz (Tashkurgan), Thailand (Lawa from Chiang Mai Province and Mae Hong Son Province,[37] Blang from Chiang Rai Province,[37] Mon from Ratchaburi Province[37]), Vietnam (Phù Lá, Hà Nhì)
- A18 – Japan, China (Han from Fujian, Han from Beijing, Han from Lanzhou[39]</ref>), Romania
- A19 – China (Han from Beijing, etc.)
- A20 – Japan, Han Chinese (Denver)
- A21 – Tibet (Sherpa, Deng, etc.), Jammu and Kashmir
- A22 – China, Han Chinese (Denver)
- A24 – China (Han in Beijing), Turkey, Czech Republic (West Bohemia)
- A25 – Japan (Chiba), China, Yakut (Vilyuy River basin)
- A26 – Denmark
- A1 [TMRCA 12,800 (95% CI 6,500 <-> 22,700) ybp[1]]
- A3 – Japan (Tokyo, etc.), Korea [TMRCA 6,800 (95% CI 3,200 <-> 12,600) ybp[1]]
- A3a – Japan (Aichi, etc.) [TMRCA 4,300 (95% CI 1,400 <-> 9,800) ybp[1]]
- A7 [TMRCA 8,800 (95% CI 5,400 <-> 13,500) ybp[1]]
- A9
- A11 – Nepal, Korea,[1] Russia [TMRCA 14,500 (95% CI 9,700 <-> 20,800) ybp[1]]
- A+T152C!+T16362C – Uyghur, Korea, Japan, Vietnam (Hmong from Lao Cai Province,[26] Kinh from Hanoi,[26] Cờ Lao)
- A5 – China (incl. Hong Kong), Japan [TMRCA 16,200 (95% CI 11,100 <-> 22,800) ybp]
- A5a – Japan (Tokyo, Aichi, etc.), Korea, China [TMRCA 5,500 (95% CI 3,800 <-> 7,600) ybp[1]]
- A5a1 - Korea
- A5a1a – Japan (Tokyo, etc.), Korea
- A5a1a1 – Japan (Tokyo, Chiba, Aichi, etc.), Korea[40]
- A5a1a1a – Japan (Tokyo, etc.)
- A5a1a1b – Japan (Tokyo, Chiba, etc.), Korea
- A5a1a2 – Japan, Korea
- A5a1a2a – Japan (Aichi)
- A5a1a1 – Japan (Tokyo, Chiba, Aichi, etc.), Korea[40]
- A5a1b – Japan (Tokyo, Aichi)
- A5a1a – Japan (Tokyo, etc.), Korea
- A5a2 – Japan (Tokyo, Aichi, etc.)
- A5a3
- A5a3* – Korea, USA (African American)
- A5a3a
- A5a3a* – Japan (Tokyo)
- A5a3a1 – Japan (Tokyo, Aichi, etc.)
- A5a4 – Japan
- A5a5 – Japan, South Korea (Seoul), Uyghur
- A5a1 - Korea
- A5b – China (Tujia, Hui, etc.) [TMRCA 12,800 ybp (95% CI 8,400 <-> 18,800) ybp[1]]
- A5b1 – China (Han from Beijing, etc.), Japan, Korea, Uyghur, Thailand, Vietnam (Tay), Singapore [TMRCA 8,600 (95% CI 6,600 <-> 11,100) ybp[1]]
- A5b1* – Uyghur
- A5b1a – Japan (Tokyo, etc.), Korea[29] [TMRCA 6,700 (95% CI 3,700 <-> 11,300) ybp[1]]
- A5b1b – China (Han from Fujian, Miao, etc.), Uyghur, Korea[41] [TMRCA 7,300 (95% CI 5,600 <-> 9,400) ybp[1]]
- A5b1b* – Han Chinese
- A5b1b1
- A5b1b1* – Miao
- A5b1b1a – China
- A5b1b1b – China
- A5b1b2 – Uyghur
- A5b1c – Han Chinese (Denver) [TMRCA 7,600 (95% CI 3,100 <-> 15,500) ybp[1]]
- A5b1c1 – Taiwan (Hakka, Bunun, Paiwan) [TMRCA 5,400 (95% CI 1,800 <-> 12,600) ybp[1]]
- A5b1d [TMRCA 7,300 (95% CI 3,700 <-> 13,000) ybp[1]]
- A5b1d* – China
- A5b1d1 – Siamese (Central Thailand), Tay (Vietnam)
- A5b2 – China (Tujia, etc.)
- A5b1 – China (Han from Beijing, etc.), Japan, Korea, Uyghur, Thailand, Vietnam (Tay), Singapore [TMRCA 8,600 (95% CI 6,600 <-> 11,100) ybp[1]]
- A5c – Japan (Aichi, etc.), Korea,[41] Khamnigan, Buryat, Barghut[1] [TMRCA 8,200 (95% CI 4,800 <-> 13,000) ybp[1]]
- A5c1 – Japan (Tokyo, Chiba, Aichi, etc.)
- A5a – Japan (Tokyo, Aichi, etc.), Korea, China [TMRCA 5,500 (95% CI 3,800 <-> 7,600) ybp[1]]
- A8 – Uyghur [TMRCA 14,000 (95% CI 9,500 <-> 19,800) ybp[1]]
- A10 – China (Uyghur), Afghanistan (Hazara, Uzbek), Russia (Mansi, Volga Tatars, etc.), France, Canada, New York [TMRCA 9,200 (95% CI 4,900 <-> 15,600) ybp[1]]
- A(xA5, A8, A10) – China (Han from Wuhan), Buryat (Inner Mongolia)
Popular culture
The mummy "Juanita" of Peru, also called the "Ice Maiden", has been shown to belong to mitochondrial haplogroup A.[42][43]
In his popular book The Seven Daughters of Eve, Bryan Sykes named the originator of this mtDNA haplogroup Aiyana.
Eva Longoria, an American actress of Mexican descent, belongs to Haplogroup A2.
See also
- Genealogical DNA test
- Genetic genealogy
- Human mitochondrial genetics
- Population genetics
- Indigenous Amerindian genetics
|
Phylogenetic tree of human mitochondrial DNA (mtDNA) haplogroups | |||||||||||||||||||||||||||||||||||||||
| Mitochondrial Eve (L) | |||||||||||||||||||||||||||||||||||||||
| L0 | L1–6 | ||||||||||||||||||||||||||||||||||||||
| L1 | L2 | L3 | L4 | L5 | L6 | ||||||||||||||||||||||||||||||||||
| M | N | ||||||||||||||||||||||||||||||||||||||
| CZ | D | E | G | Q | O | A | S | R | I | W | X | Y | |||||||||||||||||||||||||||
| C | Z | B | F | R0 | pre-JT | P | U | ||||||||||||||||||||||||||||||||
| HV | JT | K | |||||||||||||||||||||||||||||||||||||
| H | V | J | T | ||||||||||||||||||||||||||||||||||||
References
- YFull MTree 1.01.5539
- Behar et al., 2012b
- van Oven, Mannis; Manfred Kayser (13 Oct 2008). "Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation". Human Mutation. 30 (2): E386–E394. doi:10.1002/humu.20921. PMID 18853457. S2CID 27566749.
- Fagundes, Nelson J.R.; Ricardo Kanitz; Roberta Eckert; Ana C.S. Valls; Mauricio R. Bogo; Francisco M. Salzano; David Glenn Smith; Wilson A. Silva; Marco A. Zago; Andrea K. Ribeiro-dos-Santos; Sidney E.B. Santos; Maria Luiza Petzl-Erler; Sandro L.Bonatto (2008). "Mitochondrial Population Genomics Supports a Single Pre-Clovis Origin with a Coastal Route for the Peopling of the Americas" (PDF). American Journal of Human Genetics. 82 (3): 583–592. doi:10.1016/j.ajhg.2007.11.013. PMC 2427228. PMID 18313026. Archived from the original (PDF) on 2009-03-25. Retrieved 2009-11-19.
- Tanaka, Masashi; et al. (2004). "Mitochondrial Genome Variation in Eastern Asia and the Peopling of Japan". Genome Research. 14 (10A): 1832–1850. doi:10.1101/gr.2286304. PMC 524407. PMID 15466285.
- Miroslava Derenko, Boris Malyarchuk, Tomasz Grzybowski et al., "Phylogeographic Analysis of Mitochondrial DNA in Northern Asian Populations", Am. J. Hum. Genet. 2007;81:1025–1041. DOI: 10.1086/522933
- Volodko, Natalia V.; Starikovskaya, Elena B.; Mazunin, Ilya O.; Eltsov, Nikolai P.; Naidenko, Polina V.; Wallace, Douglas C.; Sukernik, Rem I. (9 May 2008). "Mitochondrial Genome Diversity in Arctic Siberians, with Particular Reference to the Evolutionary History of Beringia and Pleistocenic Peopling of the Americas". The American Journal of Human Genetics. 82 (5): 1084–1100. doi:10.1016/j.ajhg.2008.03.019. PMC 2427195. PMID 18452887.
- Ville N Pimenoff, David Comas, Jukka U Palo et al., "Northwest Siberian Khanty and Mansi in the junction of West and East Eurasian gene pools as revealed by uniparental markers", European Journal of Human Genetics (2008) 16, 1254–1264; doi:10.1038/ejhg.2008.101
- Fuku, Noriyuki; Park, Kyong Soo; Yamada, Yoshiji; Nishigaki, Yutaka; Cho, Young Min; Matsuo, Hitoshi; Segawa, Tomonori; Watanabe, Sachiro; Kato, Kimihiko; Yokoi, Kiyoshi; Nozawa, Yoshinori; Lee, Hong Kyu; Tanaka, Masashi (March 2007). "Mitochondrial Haplogroup N9a Confers Resistance against Type 2 Diabetes in Asians". The American Journal of Human Genetics. 80 (3): 407–415. doi:10.1086/512202. PMC 1821119. PMID 17273962.
- Fuyun Ji, Mark S. Sharpley, Olga Derbeneva et al., "Mitochondrial DNA variant associated with Leber hereditary optic neuropathy and high-altitude Tibetans", PNAS (May 8, 2012), vol. 109, no. 19, 7391–7396. doi: 10.1073/pnas.1202484109
- Han-Jun Jin, Chris Tyler-Smith and Wook Kim, "The Peopling of Korea Revealed by Analyses of Mitochondrial DNA and Y-Chromosomal Markers" PLoS ONE (2009)
- Qing-Peng Kong, Yong-Gang Yao, Mu Liu et al., "Mitochondrial DNA sequence polymorphisms of five ethnic populations from northern China", Hum Genet (2003) 113 : 391–405. doi:10.1007/s00439-003-1004-7
- Kazuo Umetsu, Masashi Tanaka, Isao Yuasa et al., "Multiplex amplified product-length polymorphism analysis of 36 mitochondrial single-nucleotide polymorphisms for haplogrouping of East Asian populations", Electrophoresis (2005), 26, 91–98. DOI 10.1002/elps.200406129
- Asari, Masaru; Umetsu, Kazuo; Adachi, Noboru; Azumi, Jun-ichi; Shimizu, Keiko; Shiono, Hiroshi (September 2007). "Utility of haplogroup determination for forensic mtDNA analysis in the Japanese population". Legal Medicine. 9 (5): 237–240. doi:10.1016/j.legalmed.2007.01.007. PMID 17467322.
- Zheng, Hong-Xiang; Yan, Shi; Qin, Zhen-Dong; Wang, Yi; Tan, Jing-Ze; Li, Hui; Jin, Li (6 October 2011). "Major Population Expansion of East Asians Began before Neolithic Time: Evidence of mtDNA Genomes". PLOS ONE. 6 (10): e25835. Bibcode:2011PLoSO...625835Z. doi:10.1371/journal.pone.0025835. PMC 3188578. PMID 21998705.
- Malyarchuk, B.; Derenko, M.; Denisova, G.; Kravtsova, O. (1 October 2010). "Mitogenomic Diversity in Tatars from the Volga-Ural Region of Russia". Molecular Biology and Evolution. 27 (10): 2220–2226. doi:10.1093/molbev/msq065. PMID 20457583.
- Marchani, EE; Watkins, WS; Bulayeva, K; Harpending, HC; Jorde, LB (2008). "Culture creates genetic structure in the Caucasus: Autosomal, mitochondrial, and Y-chromosomal variation in Daghestan". BMC Genetics. 9: 47. doi:10.1186/1471-2156-9-47. PMC 2488347. PMID 18637195.
- Kristina A. Tabbada, Jean Trejaut, Jun-Hun Loo et al., "Philippine Mitochondrial DNA Diversity: A Populated Viaduct between Taiwan and Indonesia?", Mol. Biol. Evol. 27(1):21–31. (2010) doi:10.1093/molbev/msp215
- Basnet, Rajdip; Rai, Niraj; Tamang, Rakesh; Awasthi, Nagendra Prasad; Pradhan, Isha; Parajuli, Pawan; Kashyap, Deepak; Reddy, Alla Govardhan; Chaubey, Gyaneshwer; Das Manandhar, Krishna; Shrestha, Tilak Ram; Thangaraj, Kumarasamy (2022-10-15). "The matrilineal ancestry of Nepali populations" (PDF). Human Genetics. 142 (2): 167–180. doi:10.1007/s00439-022-02488-z. ISSN 0340-6717. PMID 36242641. S2CID 252904281.
- Lorenz, Joseph G.; Smith, David Glenn (November 1996). "Distribution of Four Founding mtDNA Haplogroups Among Native North Americans". American Journal of Physical Anthropology. 101 (3): 311. doi:10.1002/(SICI)1096-8644(199611)101:3<307::AID-AJPA1>3.0.CO;2-W. PMID 8922178. Retrieved 2023-05-24.
- Johnson, John R.; Lorenz, Joseph G. (2006). "Genetics, Linguistics, and Prehistoric Migrations: An Analysis of California Indian MitochondrialDNA Lineages" (PDF). Journal of California and Great Basin Anthropology. 26 (1): 46–48. Retrieved 2023-05-24.
- Breschini, Gary S.; Haversat, Trudy (2004). "Ancient DNA – Modern Connections: Results of Mitochondrial DNA Analyses from Monterey County, California" (PDF). Pacific Coast Archaeological Society Quarterly. 40 (2): 1–10. Retrieved 2023-05-24.
- Two skeletons, one each from CA-MNT-1489 and CA-MNT-1931, Late Period archeological sites located in Rancho San Carlos, inland from Carmel and south of the Carmel River, were both determined to be of haplotype A01 (Breschini and Haversat 2004).[22] An adult and child, dating from Cal BP 200, buried at CA-MNT-831, a site in Pacific Grove, on the Monterey Peninsula, both belonged to haplogroup D01 (Breschini and Haversat 2004).[22] Of four Ohlone mtDNA lineages identified by Johnson and Lorenz (2006),[21] two belonged to haplogroup C, one each to haplogroups B and D, and none to haplogroup A. Of these eight Ohlone individuals, two belonged to haplogroup A.
- Dryomov, Stanislav V.; Nazhmidenova, Azhar M.; Starikovskaya, Elena B.; Shalaurova, Sofia A.; Rohland, Nadin; Mallick, Swapan; Bernardos, Rebecca; Derevianko, Anatoly P.; Reich, David; Sukernik, Rem I. (January 28, 2021). "Mitochondrial genome diversity on the Central Siberian Plateau with particular reference to the prehistory of northernmost Eurasia". PLOS ONE. 16 (1): 5. Bibcode:2021PLoSO..1644228D. doi:10.1371/journal.pone.0244228. PMC 7842996. PMID 33507977. Retrieved 2023-05-24.
- Sato, Takehiro; Amano, Tetsuya; Ono, Hiroko; Ishida, Hajime; Kodera, Haruto; Matsumura, Hirofumi; Yoneda, Minoru; Masuda, Ryuichi (2009). "Mitochondrial DNA haplogrouping of the Okhotsk people based on analysis of ancient DNA: an intermediate of gene flow from the continental Sakhalin people to the Ainu". Anthropological Science. 117 (3): 171–180. doi:10.1537/ase.081202.
- S. Pischedda, R. Barral-Arca, A. Gómez-Carballa, et al. (2017), "Phylogeographic and genome-wide investigations of Vietnam ethnic groups reveal signatures of complex historical demographic movements." Scientific Reports volume 7, Article number: 12630 (2017).
- Kristiina Tambets, Bayazit Yunusbayev, Georgi Hudjashov, et al. (2018), "Genes reveal traces of common recent demographic history for most of the Uralic-speaking populations." Genome Biology (2018) 19:139. https://doi.org/10.1186/s13059-018-1522-1
- Fernandes, D.M., Sirak, K.A., Ringbauer, H. et al. "A genetic history of the pre-contact Caribbean." Nature (2020). https://doi.org/10.1038/s41586-020-03053-2
- Pham VH, Nguyen VL, Jung HE, Cho YS, Shin JG. "The frequency of the known mitochondrial variants associated with drug-induced toxicity in a Korean population." BMC Med Genomics. 2022 Jan 3;15(1):3. doi: 10.1186/s12920-021-01153-0. PMID: 34980117; PMCID: PMC8722126.
- Duggan AT, Whitten M, Wiebe V, Crawford M, Butthof A, et al. (2013), "Investigating the Prehistory of Tungusic Peoples of Siberia and the Amur-Ussuri Region with Complete mtDNA Genome Sequences and Y-chromosomal Markers." PLoS ONE 8(12): e83570. doi:10.1371/journal.pone.0083570
- Marchi, N., Hegay, T., Mennecier, P., Georges, M., Laurent, R., Whitten, M., Endicott, P., Aldashev, A., Dorzhu, C., Nasyrova, F., Chichlo, B., Segurel, L. and Heyer, E., "Sex-specific genetic diversity is shaped by cultural factors in Inner Asian human populations." Am. J. Phys. Anthropol. (2017)
- Kilinc, G.M., Kashuba, N., Yaka, R., Sumer, A.P., Yuncu, E., Shergin, D., Ivanov, G.L., Kichigin, D., Pestereva, K., Volkov, D., Mandryka, P., Kharinskii, A., Tishkin, A., Ineshin, E., Kovychev, E., Stepanov, A., Alekseev, A., Fedoseeva, S.A., Somel, M., Jakobsson, M., Krzewinska, M., Stora, J., and Gotherstrom, A., "Investigating Holocene human population history in North Asia using ancient mitogenomes." Sci Rep 8 (1), 8969 (2018)
- Endre Neparáczki, Klaudia Kocsy, Gábor Endre Tóth, Zoltán Maróti, Tibor Kalmár, Péter Bihari, István Nagy, György Pálfi, Erika Molnár, István Raskó, and Tibor Török, "Revising mtDNA haplotypes of the ancient Hungarian conquerors with next generation sequencing." PLoS ONE 12(4): e0174886. https://doi.org/10.1371/journal.pone.0174886
- Lippold, Sebastian; Xu, Hongyang; Ko, Albert; Li, Mingkun; Renaud, Gabriel; Butthof, Anne; Schröder, Roland; Stoneking, Mark (2014). "Human paternal and maternal demographic histories: insights from high-resolution Y chromosome and mtDNA sequences". Investigative Genetics. 5 (1): 13. doi:10.1186/2041-2223-5-13. ISSN 2041-2223. PMC 4174254. PMID 25254093.
- Derenko M, Malyarchuk B, Bahmanimehr A, Denisova G, Perkova M, et al. (2013), "Complete Mitochondrial DNA Diversity in Iranians." PLoS ONE 8(11): e80673. doi:10.1371/journal.pone.0080673
- Filipa Simão, Christina Strobl, Carlos Vullo, et al., "The maternal inheritance of Alto Paraná revealed by full mitogenome sequences." FSI Genetics Volume 39, P66-72, March 01, 2019. Published online December 19, 2018. DOI:https://doi.org/10.1016/j.fsigen.2018.12.007
- Wibhu Kutanan, Jatupol Kampuansai, Metawee Srikummool, Daoroong Kangwanpong, Silvia Ghirotto, Andrea Brunelli, and Mark Stoneking, "Complete mitochondrial genomes of Thai and Lao populations indicate an ancient origin of Austroasiatic groups and demic diffusion in the spread of Tai–Kadai languages." Hum Genet 2016 DOI 10.1007/s00439-016-1742-y.
- Wibhu Kutanan, Rasmi Shoocongdej, Metawee Srikummool, et al. (2020), "Cultural variation impacts paternal and maternal genetic lineages of the Hmong-Mien and Sino-Tibetan groups from Thailand." European Journal of Human Genetics. https://doi.org/10.1038/s41431-020-0693-x
- Hongbin Yao, Mengge Wang, Xing Zou, et al., "New insights into the fine-scale history of western-eastern admixture of the northwestern Chinese population in the Hexi Corridor via genome-wide genetic legacy." Mol Genet Genomics 2021 Mar 1. doi: 10.1007/s00438-021-01767-0.
- Qing-Peng Kong, Hans-Jürgen Bandelt, Chang Sun, et al., "Updating the East Asian mtDNA phylogeny: a prerequisite for the identification of pathogenic mutations." Human Molecular Genetics, 2006, Vol. 15, No. 13 2076–2086. doi:10.1093/hmg/ddl130
- Hwan Young Lee, Ji-Eun Yoo, Myung Jin Park, Ukhee Chung, Chong-Youl Kim, and Kyoung-Jin Shin, "East Asian mtDNA haplogroup determination in Koreans: Haplogroup-level coding region SNP analysis and subhaplogroup-level control region sequence analysis." Electrophoresis (2006). DOI 10.1002/elps.200600151.
- "The peopling of the Americas: Genetic ancestry influences health". Scientific American.
- "First Americans Endured 20,000-Year Layover – Jennifer Viegas, Discovery News". Archived from the original on 2012-03-13. Retrieved 2009-11-18.
External links
- General
- Ian Logan's Mitochondrial DNA Site
- Mannis van Oven's Phylotree
- Haplogroup A
- Beringian Standstill and Spread of Native American Founders (PLoS)
- Spread of Haplogroup A, from National Geographic
- Aiyana
- A10 Ancient DNA – Ancient Chumash Paleoasiatic remains. A10 haplogroup assignment. www.pcas.org/assets/docu Results of Mitochondrial DNA Analyses from Monterey County, California
- A10 (( A10 ancient America sample.......Chumash documented origin, Cayegues (Kayiwish) Coastal Paleoasiatic California Indian populations, John R. Johnson, Anthropology 131CA http://www.anth.ucsb.edu/classes/anth131ca/California%20Genetic%20Prehistory.pdf
- A10 In the Johnson & Lorenz (2006) study, case JJ168 (haploTYPE A10), does not qualify the PhyloTree.org criteria to be classified as belonging to haploGROUP A10. In HVR1 (the only region their research tested), JJ168 does not present criterion mutations 16227c nor 16311.