Legionella pneumophila
Legionella pneumophila is a thin, aerobic, pleomorphic, flagellated, non-spore-forming, Gram-negative bacterium of the genus Legionella.[1][2] L. pneumophila is the primary human pathogenic bacterium in this group and is the causative agent of Legionnaires' disease, also known as legionellosis.
| Legionella pneumophila | |
|---|---|
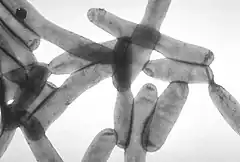 | |
| Transmission electron microscopy image of L. pneumophila | |
| Scientific classification | |
| Domain: | Bacteria |
| Phylum: | Pseudomonadota |
| Class: | Gammaproteobacteria |
| Order: | Legionellales |
| Family: | Legionellaceae |
| Genus: | Legionella |
| Species: | L. pneumophila |
| Binomial name | |
| Legionella pneumophila Brenner DJ, Steigerwalt AG, McDade JE 1979 | |
In nature, L. pneumophila infects freshwater and soil amoebae of the genera Acanthamoeba and Naegleria.[3] The mechanism of infection is similar in amoeba and human cells.
Characterization
L. pneumophila is a Gram-negative, non-encapsulated, aerobic bacillus with a single, polar flagellum often characterized as being a coccobacillus. It is aerobic and unable to hydrolyse gelatin or produce urease. It is also non-fermentative. L. pneumophila is neither pigmented nor does it autofluoresce. It is oxidase- and catalase-positive, and produces beta-lactamase. L. pneumophila colony morphology is gray-white with a textured, cut-glass appearance; it also requires cysteine and iron to thrive. It grows on yeast extract agar in "opal-like" colonies.
Cell membrane structure
While L. pneumophila is categorized as a Gram-negative organism, it stains poorly due to its unique lipopolysaccharide content in the outer leaflet of the outer cell membrane.[4] The bases for the somatic antigen specificity of this organism are located on the side chains of its cell wall. The chemical composition of these side chains both with respect to components and arrangement of the different sugars, determines the nature of the somatic or O-antigenic determinants, which are important means of serologically classifying many Gram-negative bacteria. At least 35 different serovars of L. pneumophila have been described, as well as several other species being subdivided into a number of serovars.
Detection
Sera have been used both for slide agglutination studies and for direct detection of bacteria in tissues using immunofluorescence via fluorescent-labelled antibody. Specific antibody in patients can be determined by the indirect fluorescent antibody test. ELISA and microagglutination tests have also been successfully applied.
Legionella stains poorly with Gram stain, stains positive with silver, and is cultured on charcoal yeast extract with iron and cysteine.
Ecology and reservoirs
L. pneumophila is a bacterium that can be found in numerous different environmental conditions. It can preside in temperatures ranging from 0-63 °C, a pH range of 5.0-8.5, and in dissolved oxygen levels of 0.2-15.0 mg/liter.[5]
In the environment, it can be found in freshwater environments within this wide range of temperatures. Although it can be found in this wider range, it only multiplies within a temperature of 25 and 42 °C.[5] With their ability to preside in water, they can also resist chlorination of water and pass into water control systems easily. [5] With this ability to infiltrate water systems, they can form biofilms in the walls of pipes which can lead to this bacterium being aerosolized through faucets, showers, sprinklers, and other fixtures, leading to infection after prolonged exposure.[6] With the main cause of L. pneumophila contamination being the water supply network, this has caused L. pneumophila to grow and proliferate in places such as: cooling towers, water systems of hospitals, hotels, and cruise ships. [5]
As a facultative intracellular parasite, L. pneumophila can invade and replicate inside amoebae in the environment, especially within the species of the genera Acanthamoeba and Naegleria, which can thus serve as a reservoir for L. pneumophila. These hosts will then provide protection against unfavorable physical and chemical conditions, such as chlorination.[7]
Biofilms
Biofilms are specialized, surface attachment communities that can consist of one microbe, or multiple different ones, ranging from bacteria, algae, and protozoa. Biofilms on plumbing systems and in water distribution systems is where a lot of L. pneumophila can survive. [5] Between 2009 and 2010, L. pneumophila contributed to 58% of the total waterborne disease outbreaks associated with drinking water in the United States, causing an increase in research surrounding L. pneumophila biofilms and proliferation.[8] Material also plays a role in biofilm proliferation. In water piping, L. pneumophila was more commonly found in plastic pipes at 40°C, rather than a copper pipe, which actually inhibited growth.[5]
Free-Living Protozoa
Legionella is a species that is known to infect and multiply within species of free-living amoeba. We know of at least 20 different species of free-living amoeba that support the intracellular replication of L. pneumophila. [9] This bacterium can infect and survive within the amoeba genera which includes: Acanthamoeba, Vermamoeba and Naegleria. L. pneumophila are surrounded by a amoeba-resistant cyst while residing inside the amoeba, allowing them to survive harsh environmental conditions such as chlorine, which is commonly used in water treatment systems. [10]
Although it is known that free-living amoeba play an important role in the ecology of L. pneumophila, there is little data on how these amoebas interact with other amoebas, and how this affects L. pneumophila overall. [9]
Frequency of occurrence
L. pneumopila is the primary (>90%) causative organism for Legionnaires disease.[11] Approximately 1.9 per 100,000 people are infected with Legionnaires disease each year in the EU.[12] L. pneumopila often infects individuals through poor quality water sources. Approximately 20% of reported Legionnaires disease cases come from healthcare, senior living, or travel facilities that have been exposed to water contaminated with L. pneumopila[11].
Pathogenesis
In humans, L. pneumophila invades and replicates inside macrophages. The internalization of the bacteria can be enhanced by the presence of antibody and complement, but is not absolutely required. Internalization of the bacteria appears to occur through phagocytosis. However, L. pneumophila is also capable of infecting non-phagocytic cells through an unknown mechanism. A rare form of phagocytosis known as coiling phagocytosis has been described for L. pneumophila, but this is not dependent on the Dot/Icm (intracellular multiplication/defect in organelle trafficking genes) bacterial secretion system and has been observed for other pathogens.[13] Once internalized, the bacteria surround themselves in a membrane-bound vacuole that does not fuse with lysosomes that would otherwise degrade the bacteria. In this protected compartment, the bacteria multiply.
Dot/Icm type IV secretion system
The bacteria use a type IVB secretion system known as Dot/Icm to inject effector proteins into the host. These effectors are involved in increasing the bacteria's ability to survive inside the host cell. L. pneumophila encodes for over 330 "effector" proteins,[14] which are secreted by the Dot/Icm translocation system to interfere with host cell processes to aid bacterial survival. It has been predicted that the genus Legionella encodes more than 10,000 and possibly up to ~18,000 effectors that have a high probability to be secreted into their host cells.[15][16]
One main way in which L. pneumophila uses its effector proteins is to interfere with fusion of the Legionella-containing vacuole with the host's endosomes, and thus protect against lysis.[17] Knock-out studies of Dot/Icm translocated effectors indicate that they are vital for the intracellular survival of the bacterium, but many individual effector proteins are thought to function redundantly, in that single-effector knock-outs rarely impede intracellular survival. This high number of translocated effector proteins and their redundancy is likely a result of the bacterium having evolved in many different protozoan hosts.[18]
Legionella-containing vacuole

For Legionella to survive within macrophages and protozoa, it must create a specialized compartment known as the Legionella-containing vacuole (LCV). Through the action of the Dot/Icm secretion system, the bacteria are able to prevent degradation by the normal endosomal trafficking pathway and instead replicate. Shortly after internalization, the bacteria specifically recruit endoplasmic reticulum-derived vesicles and mitochondria to the LCV while preventing the recruitment of endosomal markers such as Rab5a and Rab7a. Formation and maintenance of the vacuoles are crucial for pathogenesis; bacteria lacking the Dot/Icm secretion system are not pathogenic and cannot replicate within cells, while deletion of the Dot/Icm effector SdhA results in destabilization of the vacuolar membrane and no bacterial replication.[19][20]
Nutrient acquisition
Legionella is auxotrophic for seven amino acids: cysteine, leucine, methionine, valine, threonine, isoleucine, and arginine. Once inside the host cell, Legionella needs nutrients to grow and reproduce. Inside the vacuole, nutrient availability is low; the high demand of amino acids is not covered by the transport of free amino acids found in the host cytoplasm. To improve the availability of amino acids, the parasite promotes the host mechanisms of proteasomal degradation. This process in L. pneumophila includes the SCF1 ubiquitin ligase and the AnkB F-Box effector, which is farnesylated by the activity of three host enzymes localized in the membrane of the LCV: farnesyltransferase, Ras-converting enzyme-1 protease, and ICMT. Farnesylation allows AnkB to get anchored into the cytoplasmic side of the vacuole. SCF1 and AnkB interact with each other to degrade Lys-linked polyubiquitinated proteins.[21] This generates an excess of free amino acids in the cytoplasm of L. pneumophila-infected cells that can be used for intravacuolar proliferation of the parasite.
The K48-linked polyubiquitination is a marker for proteasomal degradation that releases 2 to 24-amino-acid-long peptides, which are quickly degraded to amino acids by various oligopeptidases and aminopeptidases present in the cytoplasm. Amino acids are imported into the LCV through various amino acid transporters such as the neutral amino acid transporter B(0).[22]
The amino acids are the primary carbon and energy source of L. pneumophila, that have almost 12 classes of ABC-transporters, amino acid permeases, and many proteases, to exploit it. The imported amino acids are used by L. pneumophila to generate energy through the TCA cycle (Krebs cycle) and as sources of carbon and nitrogen. Because the amino acid degradation acts as the main carbon source for L. pneumophila, this microbe has a lack of requirement for glucose. Despite this, L. pneumophila does contain multiple amylases, such as LamB, which hydrolyzes polysaccharides into glucose monomers for metabolism. The loss of LamB can result in severe growth issues for L. pneumophila.[23]
However, promotion of proteasomal degradation for the obtention of amino acids and the hydrolyzation of polysaccharides may not be the only virulence strategies to obtain carbon and energy sources from the host. Type II–secreted degradative enzymes may provide an additional strategy to generate carbon and energy sources.[24]
L. pneumophila is the only known intracellular pathogen to have a Type II Secretion System (secretome). In Type II Secretion, proteins are first translocated across the inner membrane into the periplasmic space. This process is mediated by either the Sec or Tat pathway. Soon after, the same proteins are then transported through a specific pore in the outer membrane to the exterior of the cell. This secretome is believed to have as many as 60 proteins incorporated into the system. [24]
Genomics
| NCBI genome ID | 416 |
|---|---|
| Ploidy | haploid |
| Genome size | 3.44 Mb |
| Number of chromosomes | 1 |
| Year of completion | 2004 |
The determination and publication of the complete genome sequences of three clinical L. pneumophila isolates in 2004 paved the way for the understanding of the molecular biology of L. pneumophila in particular and Legionella in general. In-depth comparative genome analysis using DNA arrays to study the gene content of 180 Legionella strains revealed high genome plasticity and frequent horizontal gene transfer. Further insight in the L. pneumophila lifecycle was gained by investigating the gene expression profile of L. pneumophila in Acanthamoeba castellanii, its natural host. L. pneumophila exhibits a biphasic lifecycle and defines transmissive and replicative traits according to gene expression profiles.[2]
Genetic transformation
Transformation is a bacterial adaptation involving the transfer of DNA from one bacterium to another through the surrounding liquid medium. Transformation is a bacterial form of sexual reproduction.[25] In order for a bacterium to bind, take up, and recombine exogenous DNA into its chromosome, it must enter a special physiological state referred to as "competence".
To determine which molecules may induce competence in L. pneumophila, 64 toxic molecules were tested.[26] Only six of these molecules, all DNA-damaging agents, caused strong induction of competence. These were mitomycin C (which introduces DNA inter-strand crosslinks), norfloxacin, ofloxacin, and nalidixic acid (inhibitors of DNA gyrase that cause double-strand breaks), bicyclomycin (causes double-strand breaks), and hydroxyurea (causes oxidation of DNA bases). These results suggest that competence for transformation in L. pneumophila evolved as a response to DNA damage.[26] Perhaps induction of competence provides a survival advantage in a natural host, as occurs with other pathogenic bacteria.[25]
Drug targets
Several enzymes in the bacteria have been proposed as tentative drug targets. For example, enzymes in the iron uptake pathway have been suggested as important drug targets.[27] Further, a cN-II class of IMP/GMP specific 5´-nucleotidase which has been extensively characterized kinetically. The tetrameric enzyme shows aspects of positive homotropic cooperativity, substrate activation and presents a unique allosteric site that can be targeted to design effective drugs against the enzyme and thus, the organism. Moreover, the enzyme is distinct than its human counterpart making it an attractive target for drug development.
Treatment
Macrolides (azithromycin or clarithromycin) or fluoroquinolones (levofloxacin or moxifloxacin) are the standard treatment for Legionella pneumonia in humans, with levofloxacin being considered first line with increasing resistance to azithromycin. Two studies support superiority of levofloxacin over macrolides, although not FDA approved.[28]
References
- Madigan M, Martinko J, eds. (2005). Brock Biology of Microorganisms (11th ed.). Prentice Hall. ISBN 0-13-144329-1.
- Heuner K, Swanson M, eds. (2008). Legionella: Molecular Microbiology. Caister Academic Press. ISBN 978-1-904455-26-4.
- Rowbotham TJ (December 1980). "Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae". Journal of Clinical Pathology. 33 (12): 1179–1183. doi:10.1136/jcp.33.12.1179. PMC 1146371. PMID 7451664.
- Ryan KJ, Ray CG, eds. (2004). "Chapter 26: Legionella". Sherris Medical Microbiology (4th ed.). McGraw Hill. ISBN 978-0-8385-8529-0.
- Uzel A, Hames-Kocabas EE (2010). Legionella Pneumophila: From Environment to Disease. NOVA Science Publishers. pp. 5–8. ISBN 978-1-60876-947-6.
- Abdel-Nour M, Duncan C, Low DE, Guyard C (October 2013). "Biofilms: the stronghold of Legionella pneumophila". International Journal of Molecular Sciences. 14 (11): 21660–21675. doi:10.3390/ijms141121660. PMC 3856027. PMID 24185913.
- Rowbotham TJ (December 1980). "Preliminary report on the pathogenicity of Legionella pneumophila for freshwater and soil amoebae". Journal of Clinical Pathology. 33 (12): 1179–83. doi:10.1136/jcp.33.12.1179. PMC 1146371. PMID 7451664.
- Shen Y, Monroy GL, Derlon N, Janjaroen D, Huang C, Morgenroth E, et al. (April 2015). "Role of biofilm roughness and hydrodynamic conditions in Legionella pneumophila adhesion to and detachment from simulated drinking water biofilms". Environmental Science & Technology. 49 (7): 4274–4282. Bibcode:2015EnST...49.4274S. doi:10.1021/es505842v. PMC 4472476. PMID 25699403.
- Dey R, Mameri MR, Trajkovic-Bodennec S, Bodennec J, Pernin P (September 2020). "Impact of inter-amoebic phagocytosis on the L. pneumophila growth". FEMS Microbiology Letters. 367 (18). doi:10.1093/femsle/fnaa147. PMID 32860684.
- Muchesa P, Leifels M, Jurzik L, Barnard TG, Bartie C (2018). "Detection of amoeba-associated Legionella pneumophila in hospital water networks of Johannesburg". Southern African Journal of Infectious Diseases. 33 (3): 72–75. doi:10.1080/23120053.2018.1434060. S2CID 90027194 – via SAJID.
- Donohue MJ, Pham M, Brown S, Easwaran KM, Vesper S, Mistry JH (June 2023). "Water quality influences Legionella pneumophila determination". Water Research. 238: 119989. Bibcode:2023WatRe.23819989D. doi:10.1016/j.watres.2023.119989. PMC 10351031. PMID 37137207.
- Lupia T, Corcione S, Shbaklo N, Rizzello B, De Benedetto I, Concialdi E, et al. (February 2023). "Legionella pneumophila Infections during a 7-Year Retrospective Analysis (2016-2022): Epidemiological, Clinical Features and Outcomes in Patients with Legionnaires' Disease". Microorganisms. 11 (2): 498. doi:10.3390/microorganisms11020498. PMC 9965988. PMID 36838463.
- Rittig MG, Krause A, Häupl T, Schaible UE, Modolell M, Kramer MD, et al. (October 1992). "Coiling phagocytosis is the preferential phagocytic mechanism for Borrelia burgdorferi". Infection and Immunity. 60 (10): 4205–4212. doi:10.1128/iai.60.10.4205-4212.1992. PMC 257454. PMID 1398932.
- Ensminger AW (February 2016). "Legionella pneumophila, armed to the hilt: justifying the largest arsenal of effectors in the bacterial world". Current Opinion in Microbiology. 29: 74–80. doi:10.1016/j.mib.2015.11.002. PMID 26709975.
- Burstein D, Amaro F, Zusman T, Lifshitz Z, Cohen O, Gilbert JA, et al. (February 2016). "Genomic analysis of 38 Legionella species identifies large and diverse effector repertoires". Nature Genetics. 48 (2): 167–175. doi:10.1038/ng.3481. PMC 5050043. PMID 26752266.
- Gomez-Valero L, Rusniok C, Carson D, Mondino S, Pérez-Cobas AE, Rolando M, et al. (February 2019). "More than 18,000 effectors in the Legionella genus genome provide multiple, independent combinations for replication in human cells". Proceedings of the National Academy of Sciences of the United States of America. 116 (6): 2265–2273. Bibcode:2019PNAS..116.2265G. doi:10.1073/pnas.1808016116. PMC 6369783. PMID 30659146.
- Pan X, Lührmann A, Satoh A, Laskowski-Arce MA, Roy CR (June 2008). "Ankyrin repeat proteins comprise a diverse family of bacterial type IV effectors". Science. 320 (5883): 1651–1654. Bibcode:2008Sci...320.1651P. doi:10.1126/science.1158160. PMC 2514061. PMID 18566289.
- Jules M, Buchrieser C (June 2007). "Legionella pneumophila adaptation to intracellular life and the host response: clues from genomics and transcriptomics". FEBS Letters. 581 (15): 2829–2838. doi:10.1016/j.febslet.2007.05.026. PMID 17531986. S2CID 23203471.
- Harding CR, Stoneham CA, Schuelein R, Newton H, Oates CV, Hartland EL, et al. (July 2013). "The Dot/Icm effector SdhA is necessary for virulence of Legionella pneumophila in Galleria mellonella and A/J mice". Infection and Immunity. 81 (7): 2598–2605. doi:10.1128/IAI.00296-13. PMC 3697626. PMID 23649096.
- Creasey EA, Isberg RR (February 2012). "The protein SdhA maintains the integrity of the Legionella-containing vacuole". Proceedings of the National Academy of Sciences of the United States of America. 109 (9): 3481–3486. doi:10.1073/pnas.1121286109. PMC 3295292. PMID 22308473.
- Price CT, Richards AM, Abu Kwaik Y (2014-08-26). "Nutrient generation and retrieval from the host cell cytosol by intra-vacuolar Legionella pneumophila". Frontiers in Cellular and Infection Microbiology. 4: 111. doi:10.3389/fcimb.2014.00111. PMC 4143614. PMID 25207263.
- Price CT, Richards AM, Abu Kwaik Y (2014-08-26). "Nutrient generation and retrieval from the host cell cytosol by intra-vacuolar Legionella pneumophila". Frontiers in Cellular and Infection Microbiology. 4: 111. doi:10.3389/fcimb.2014.00111. PMC 4143614. PMID 25207263.
- Best A, Price C, Ozanic M, Santic M, Jones S, Abu Kwaik Y (April 2018). "A Legionella pneumophila amylase is essential for intracellular replication in human macrophages and amoebae". Scientific Reports. 8 (1): 6340. Bibcode:2018NatSR...8.6340B. doi:10.1038/s41598-018-24724-1. PMC 5910436. PMID 29679057.
- Mintz CS (December 1999). "Gene transfer in Legionella pneumophila". Microbes and Infection. 1 (14): 1203–1209. doi:10.1016/s1286-4579(99)00241-5. PMID 10580276.
- Michod RE, Bernstein H, Nedelcu AM (May 2008). "Adaptive value of sex in microbial pathogens". Infection, Genetics and Evolution. 8 (3): 267–285. doi:10.1016/j.meegid.2008.01.002. PMID 18295550.
- Charpentier X, Kay E, Schneider D, Shuman HA (March 2011). "Antibiotics and UV radiation induce competence for natural transformation in Legionella pneumophila". Journal of Bacteriology. 193 (5): 1114–1121. doi:10.1128/JB.01146-10. PMC 3067580. PMID 21169481.
- Cianciotto NP (May 2015). "An update on iron acquisition by Legionella pneumophila: new pathways for siderophore uptake and ferric iron reduction". Future Microbiology. 10 (5): 841–851. doi:10.2217/fmb.15.21. PMC 4461365. PMID 26000653.
- Gilbert DN, Moellering Jr RC, Eliopoulos GM, Chambers HF, Saag MS, eds. (2013). The Sanford Guide to Antimicrobial Therapy (43rd ed.). Antimicrobial Therapy. ISBN 978-1-930808-75-1.