Birds: Neural Mechanism for Song Learning in Zebra Finches
Introduction
Over the past four decades songbirds have become a widely used model organism for neuroscientists studying complex sequential behaviours and sensory-guided motor learning. Like human babies, young songbirds learn many of the sounds they use for communication by imitating adults. One songbird in particular, the zebra finch (Taeniopygia guttata), has been the focus of much research because of its proclivity to sing and breed in captivity and its rapid maturation. The song of an adult male zebra finch is a stereotyped series of acoustic signals with structure and modulation over a wide range of time scales, from milliseconds to several seconds. The adult zebra finch song comprises a repeated sequence of sounds, called a motif, which lasts about a second. The motif is composed of shorter bursts of sound called syllables, which often contain sequences of simpler acoustic elements called notes as shown in Fig.1. The songbirds learning system is a very good model to study the sensory-motor integration because the juvenile bird actively listens to the tutor and modulates its own song by correcting for errors in the pitch and offset. The neural mechanism and the architecture of the song bird brain which plays a crucial role in learning is similar to the language processing region in frontal cortex of humans. Detailed study of the hierarchical neural network involved in the learning process could provide significant insights into the neural mechanism of speech learning in humans.
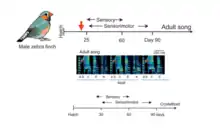
Illustration of the typical song structure & learning phases involved in song bird.
Song-learning proceeds through a series of stages, beginning with sensory phase where the juvenile bird just listens to its tutor (usually its father) vocalizing, often without producing any song-like vocalization itself. The bird uses this phase to memorize a certain structure of the tutor song, forming the neural template of the song. Then it enters the sensorimotor phase, where it starts babbling the song and correcting its errors using auditory feedback. The earliest attempt to recreate the template of the tutor song is highly noisy, unstructured and variable and it is called sub-song. An example is shown in the spectrogram in Fig.1. Through the subsequent days the bird enters a “plastic phase” where there is a significant amount of plasticity in the neural network responsible for generating highly structured syllables and the variability is reduced in the song. By the time they reach sexual maturity, the variability is substantially eliminated—a process called crystallization—and the young bird begins to produce a normal adult song, which can be a striking imitation of the tutor song (Fig.1). Thus, the gradual reduction of song variability from early sub-song to adult song, together with the gradual increase in imitation quality, is an integral aspect of vocal learning in the songbird. In the following sections we will explore several parts of the avian brain and the underlying neural mechanisms that are responsible for this remarkable vocal imitation observed in these birds.
Hierarchical Neural Network involved in the generation of song sequences
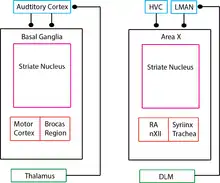
It is important to understand the neuroanatomy of the songbird in detail because it provides significant information about the learning mechanisms involved in various motor and sensory integration pathways. This could ultimately shed light on the language processing and vocal learning in humans. The exact neuroanatomical data about human speech processing system is still unknown and songbird anatomy and physiology will enable us to make plausible hypotheses. The comparison of the mammalian brain and a songbird (avian) brain is made in the final section of this chapter in (Fig. 6). The pathway observed in the avian brain can be broadly divided into motor control and anterior forebrain pathway as shown in (Fig.2). The auditory pathway provides the error feedback signals which leads to potentiation or depression of the synaptic connections involved in motor pathways, which plays a significant role in vocal learning. The motor control pathway includes Hyperstriatum Ventrale, pars Caudalis (HVC), Robust Nucleus of Acropallium (RA), Tracheosyringeal subdivision of the hypoglossal nucleus (nXIIts) and Syrinx. This pathway is necessary for generating the required motor control signals which produce highly structured songs and coordinating breathing with singing. The anterior forebrain pathway includes Lateral magnocellular nucleus of anterior nidopallium (LMAN), Area X (X) and the medial nucleus of dorsolateral thalamus (DLM). This pathway plays a crucial role in song learning in juveniles, song variability in adults and song representation. The auditory pathway includes substantia nigra (SNc) and the ventral tegmental area (VTA), which plays a crucial role in auditory inputs processing and analyzing the feedback error. The muscles of the syrinx are innervated by a subset of motor neurons from nXIIts. A primary projection to the nXIIts descends from neurons in the forebrain nucleus RA. Nucleus RA receives motor-related projections from another cortical analogue, nucleus HVC, which in turn receives direct input from several brain areas, including thalamic nucleus uvaeformis (Uva).
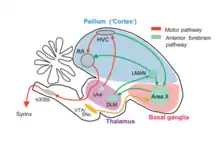
Neural Mechanism for the generation of highly structured & temporally precise syllable pattern
Nuclei HVC and RA are involved in the motor control of song in a hierarchical manner (Yu and Margoliash 1996). Recordings in singing zebra finches have shown that HVC neurons that project to RA transmit an extremely sparse pattern of bursts: each RA-projecting HVC neuron generates a single highly stereotyped burst of approximately 6 ms duration at one specific time in the song (Hahnloser, Kozhevnikov et al. 2002). During singing, RA neurons generate a complex sequence of high-frequency bursts of spikes, the pattern of which is precisely reproduced each time the bird sings its song motif (Yu and Margoliash 1996). During a motif, each RA neuron produces a fairly unique pattern of roughly 12 bursts, each lasting ~10 ms (Leonardo and Fee 2005). Based on the observations that RA-projecting HVC neurons generate a single burst of spikes during the song motif and that different neurons appear to burst at many different times in the motif, it has been hypothesized that these neurons generate a continuous sequence of activity over time (Fee, Kozhevnikov et al. 2004, Kozhevnikov and Fee 2007). In other words, at each moment in the song, there is a small ensemble of HVC (RA) neurons active at that time and only at that time (Figure 3), and each ensemble transiently activates (for ~10 ms) a subset of RA neurons determined by the synaptic connections of HVC neurons in RA (Leonardo and Fee 2005). Further, in this model the vector of muscle activities, and thus the configuration of the vocal organ, is determined by the convergent input from RA neurons on a short time scale, of about 10 to 20 ms. The view that RA neurons may simply contribute transiently, with some effective weight, to the activity of vocal muscles is consistent with some models of cortical control of arm movement in primates (Todorov 2000). A number of studies suggest that the timing of the song is controlled on a millisecond-by-millisecond basis by a wave, or chain, of activity that propagates sparsely through HVC neurons. This hypothesis is supported by an analysis of timing variability during natural singing (Glaze and Troyer 2007) as well as experiments in which circuit dynamics in HVC were manipulated to observe the effect on song timing. Thus, in this model, song timing is controlled by propagation of activity through a chain in HVC; the generic sequential activation of this HVC chain is translated, by the HVC connections in RA, into a specific precise sequence of vocal configurations.
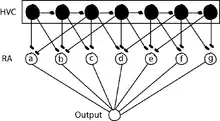
Synaptic Plasticity in Posterior Forebrain Pathway is a potential substrate for vocal learning
A number of song-related avian brain areas have been discovered (Fig. 4A). Song production areas include HVC (Hyperstriatum Ventrale, pars Caudalis) and RA (robust nucleus of the arcopallium), which generate sequences of neural activity patterns and through motor neurons control the muscles of the vocal apparatus during song (Yu and Margoliash 1996, Hahnloser, Kozhevnikov et al. 2002, Suthers and Margoliash 2002). Lesion of HVC or RA causes immediate loss of song (Vicario and Nottebohm 1988). Other areas in the anterior forebrain pathway (AFP) appear to be important for song learning but not production, at least in adults. The AFP is regarded as an avian homologue of the mammalian basal ganglia thalamocortical loop (Farries 2004). In particular, lesion of area LMAN (lateral magnocellular nucleus of the nidopallium) has little immediate effect on song production in adults, but arrests song learning in juveniles (Doupe 1993, Brainard and Doupe 2000). These facts suggest that LMAN plays a role in driving song learning, but the locus of plasticity is in brain areas related to song production, such as HVC and RA. Doya and Senjowski in 1998 proposed a tripartite schema, in which learning is based on the interactions between actor and a critic (Fig.4B). The critic evaluates the performance of the actor at a desired task. The actor uses this evaluation to change in a way that improves its performance. To learn by trial and error, the actor performs the task differently each time. It generates both good and bad variations, and the critic’s evaluation is used to reinforce the good ones. Ordinarily it is assumed that the actor generates variations by itself. However, the source of variation is external to the actor. We will call this source the experimenter. The actor was identified with HVC, RA, and the motor neurons that control vocalization. The actor learns through plasticity at the synapses from HVC to RA (Fig. 4C). Based on evidence of structural changes like axonal growth and retraction that take place in the HVC to RA projection during song learning, this view is widely regarded as a plausible mechanism. For the experimenter & critic, Doya and Senjowski turned to the anterior forebrain pathway, hypothesizing that the critic is Area X and the experimenter is LMAN.
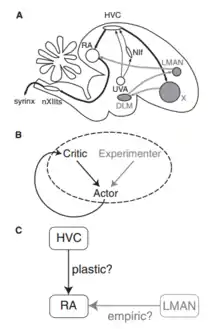
Biophysically realistic synaptic plasticity rules underlying song learning mechanism
|
Biophysically realistic model
The positive parameter , called the learning rate, controls the overall amplitude of synaptic changes. The eligibility trace is a hypothetical quantity present at every plastic synapse. It signifies whether the synapse is "eligible" for modification by reinforcement and is based on the recent activation of the plastic synapse and the empiric synapse onto the same RA neuron Here is the conductance of the empiric (LMAN->RA) synapse onto the RA neuron. The temporal filter G(t) is assumed to be nonnegative, and its shape determines how far back in time the eligibility trace can "remember" the past. The instantaneous activation of the empirical synapses is dependent on the average activity . The learning principles follows two basic rules shown in (Fig.5). First rule: If coincident activation of a plastic (HVC->RA) synapse and empiric (LMAN->RA) synapse onto the same RA neuron is followed by positive reinforcement, then the plastic synapse is strengthened. Second rule: If activation of a plastic synapse without activation of the empiric synapse onto the same RA neuron is followed by positive reinforcement, then the plastic synapse is weakened. The rules based on dynamic conductance perturbations of the actor neurons perform stochastic gradient ascent on the expected value of the reinforcement signal. This means that song performance as evaluated by the critic is guaranteed to improve on average.
|






![{\displaystyle {{e}_{ij}}(t)=\int \limits _{0}^{t}{d{t}'}\,G(t-{t}')\left[S_{i}^{LMAN}(t)-\left\langle S_{i}^{LMAN}\right\rangle \right]\,\,S_{ij}^{HVC}(t)}](../../I/b43f62b81c13062636c5d924a6ca0aa129f13fd8.svg)


Comparison between Mammalian & Songbird brain architecture
The avian Area X is homologous to the mammalian basal ganglia (BG) and includes striatal and pallidal cell types. The BG forms part of a highly conserved anatomical loop-through several stations, from cortex to the BG (striatum and pallidum), then to thalamus and back to cortex. Similar loops are seen in the songbird: the cortical analogue nucleus LMAN projects to Area X, the striatal components of which project to the thalamic nucleus DLM, which projects back to LMAN. Striatal components accounts for reward basing learning and reinforcement learning. The neuron types and its functionality are exactly comparable in Area X of birds to basal ganglia in humans as shown (in Fig.6). The close anatomical similarity motivates us to learn the song bird brain in more detail because with this we can finally achieve some significant understanding of the speech learning in humans and treat many speech related disorders with higher precision.
References
Brainard, M. S. and A. J. Doupe (2000). "Auditory feedback in learning and maintenance of vocal behaviour." Nat Rev Neurosci 1(1): 31-40.
Dembo, A. and T. Kailath (1990). "Model-free distributed learning." IEEE Trans Neural Netw 1(1): 58-70.
Doupe, A. J. (1993). "A neural circuit specialized for vocal learning." Curr Opin Neurobiol 3(1): 104-111.
Farries, M. A. (2004). "The avian song system in comparative perspective." Ann N Y Acad Sci 1016: 61-76.
Fee, M. S., A. A. Kozhevnikov and R. H. Hahnloser (2004). "Neural mechanisms of vocal sequence generation in the songbird." Ann N Y Acad Sci 1016: 153-170.Glaze, C. M. and T. W. Troyer (2007). "Behavioral measurements of a temporally precise motor code for birdsong." J Neurosci 27(29): 7631-7639.
Hahnloser, R. H., A. A. Kozhevnikov and M. S. Fee (2002). "An ultra-sparse code underlies the generation of neural sequences in a songbird." Nature 419(6902): 65-70.
Kozhevnikov, A. A. and M. S. Fee (2007). "Singing-related activity of identified HVC neurons in the zebra finch." J Neurophysiol 97(6): 4271-4283.
Leonardo, A. and M. S. Fee (2005). "Ensemble coding of vocal control in birdsong." J Neurosci 25(3): 652-661.
Seung, H. S. (2003). "Learning in spiking neural networks by reinforcement of stochastic synaptic transmission." Neuron 40(6): 1063-1073.
Suthers, R. A. and D. Margoliash (2002). "Motor control of birdsong." Curr Opin Neurobiol 12(6): 684-690.
Todorov, E. (2000). "Direct cortical control of muscle activation in voluntary arm movements: a model." Nat Neurosci 3(4): 391-398.
Vicario, D. S. and F. Nottebohm (1988). "Organization of the zebra finch song control system: I. Representation of syringeal muscles in the hypoglossal nucleus." J Comp Neurol 271(3): 346-354.
Yu, A. C. and D. Margoliash (1996). "Temporal hierarchical control of singing in birds." Science 273(5283): 1871-1875.
Birds: Magnetoperception
Introduction
Sensory magneto-perception is defined as the sense that allows an organism to detect the Earth's magnetic field and to orient itself according to it. Magneto-perception is present throughout the Bacteria and Animalia kingdoms being observed, among others, in honey bees, salamanders, fishes and frogs. Here we will explain and review the current hypotheses on how birds use the Earth's magnetic field to navigate.
During the last decade many laboratories have focused their attention on how aves (i.e. birds) orient themselves. Twice each year, migratory birds travel thousands of kilometers from their breeding region to the overwintering sites and back, finding their way even across unfamiliar territories.
Studies done especially with robins (Erithacus rubecula) and pigeons (Columba livia) have shown that in addition to inclination, intensity and polarity of the geomagnetic field, birds relies on cues such as the sun or the star map to orient. However, the importance of each cue to avian orientation is still debated.


While those studies have clarified the main aspects of birds' navigation, there are still many unanswered questions. Here we give a short introduction to magnetic orientation in animals, explaining the physics behind it, and discuss some of the principal hypotheses on how birds sense the geomagnetic field. The sensory structure, the neuronal circuitry and its mechanisms will be discussed.
Magnetic orientation
Since man cannot consciously sense the geomagnetic field, sensory magneto-reception might appear alien to our understanding. Nonetheless, the ability to sense magnetic fields is common to many animals; among them are mollusks, arthropods and members of all major group of vertebrates . The term magnetic orientation is regarded to the use these animals make of the information coming by a prominent magnetic field, the geomagnetic one, to orient themselves with respect to the Earth in migratory patterns. In this section the Earth's intrinsic magnetic field will be discussed, highlighting the two main classes of information that animals, and birds in particular, may obtain from it.
The geomagnetic field
In a first approximation, the Earth can be seen as a gigantic magnet dipole, with its poles situated close to the geographic, or rotational, poles. Although the magnetic north pole (Nm in the Figure) coincides today with the rotational north pole (Ng), there is no relationship between the two, since the latter is fixed, while the former can change in time.
An intuitive way to visualize magnetic fields is to consider their field lines. They define the direction of a vector field at different points. In a dipole (the arguably most elementary magnet) the northern and southern poles are the sources of the field. Because of the presented dipole approximation, the field lines associated with the geomagnetic field originate from the southern (magnetic, from now on) pole, run around the globe and reach the northern pole. A schematic visualization of the lines is depicted in Figure [fig:field-lines].
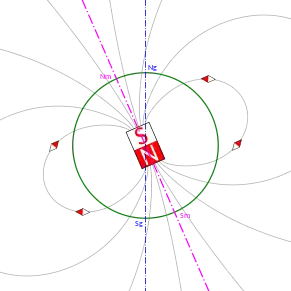


The most important aspect to note for the following discussion is that for this reason magnetic field lines point upward on the southern hemisphere, downward on the northern one, while running parallel to the Earth's surface at the magnetic equator (inclined, as the magnetic dipole, about 10 degrees with respect to the geographic equator), showing a fairly regular gradient. The intensity of the field is highest at the poles and lowest at the magnetic equator.
Of course irregularities in the Earth's surface slightly vary the real intensity of the field at different points and the corresponding inclination of its lines. Since these effects are very small, the geomagnetic field represents a reliable and omnipresent source of navigational information. In addition to the orientation of the magnetic field, which acts like a (biological) compass (as is done in human-built tools), the intensity of the field together with the inclination of the associated field line at different points may provide components of a navigational "map" indicating one's position on the globe [1].
Magnetic compass orientation
A magnetic field can be used as the main source of information to build a magnetic compass. That animals use a biological magnetic compass has been shown in several experiments, mostly involving European robins, Erithacus rubecula. A given migration pattern is in fact chosen by birds, that presents a magnetic characterization which is constant in time. Indeed recreating those fields and just inverting the poles led to analogous behaviors, inverted in direction [2].
What is most interesting is that while human-built compasses are polarity-based, featuring an orientation information based on the polarity (north/south) of the field lines, birds demonstrated to have an inclination-based compass. The gradient above described of inclinations of the filed lines, from the southern pole to the northern one, passing through the magnetic equator, can be used to detect the position of a given magnetic pole. Surprisingly, birds are not actually able to detect the full inclination of a field vector at a given point, but only of its axial component. The vertical component is inferred by simply realizing their up/down flying asset. This result has been obtained with smartly tuned magnetic fields where same axial component and different polarities led to the same outcome, birds not being able to tell the difference [3].
Another interesting aspect of birds' biological compass is that it is strictly tuned to only detect tight windows of certain magnetic fields intensities. Even more interestingly, this window may change but not in a shifting nor in an amplifying fashion. It was indeed observed that only already experienced (and orientation-wise efficient) fields are eligible as future recognizable windows Wiltschko, W. and Wiltschko, R. (1978). Further analysis of the magnetic compass of migratory birds.. Springer. 302-310.</ref>. .
Magnetic navigation
A biological compass may be enough to direct navigation, exactly as a human-built polarity-based compass is enough to orient. However, even the earliest experiments [4] showed that birds also utilize information on field intensity. Apparently discording theories have then developed throughout time suggesting that either one or the other approach was actually used by birds for navigation and orientation. Currently, it is generally accepted that the two approaches are both valid, one being prominent over the other under different conditions.
In fact, birds know by experience that in the northern hemisphere the geomagnetic field increases towards north. The difference in intensity between an encountered location and a known one lets them infer whether they are north or south of the known site. The first experimental results suggesting this ability of birds were obtained with pigeons, Columba livia f. domestica
[5] .
This is not the only way intensity can be used, though. It could also be used as a "sign-post" [1]. Birds, in fact, may show innate responses, both behavioral
Wiltschko, W. and Wiltschko, R. (1992). "Migratory orientation: magnetic compass orientation of garden warblers (sylvia borin) after a simulated crossing of the magnetic equator.". Ethology 91(1): 70-74.</ref>. and physiological , to locations that present a specific combination of field intensity and inclination of the corresponding field line. In an experiment thrush nightingales, L. luscinia, showed extremely rapid changes in weight that correlated to the geomagnetic conditions at which they originated.
Implications
In summary, inclination and intensity are both valid mechanisms that help birds in orientation and navigation. Being so different one from the other, it is clear that no single receptor or sensory system in general might be able to perceive, encode and elaborate the information these two elements represent. This is also the main reason why parallel research theories led to apparently different results when trying to solve the questions about birds' ability to orient themselves.
Magnetic sensory system
Lorenzini ampullae in fishes are electrically sensitive and specialized organs. Previous research investigated a corresponding specialized organ responsible for magnetic detection in birds. However, as it soon turned out birds' detection of the geomagnetic field is more complex than expected and does not rely exclusively on a specialized cell. The difficulty in identifying an underlying physiological mechanisms and a magneto-receptive organ or molecule has been a great obstacle in the study of the avian magnetic perception field.
There are mainly two strong hypotheses on magnetic sensing that are widely accepted in the field and strongly supported by data:
- Trigeminal iron-mineral based magneto-reception in the upper beak;
- Chemical light-dependent radical-pair based magneto-reception.
Very recently a third hypothesis has been proposed:
Inner ear lagena based magneto-reception.
The review and description of these three hypotheses is the content of the following sections.
Iron-based magneto-reception
The first proposed hypothesis on how birds sense the geomagnetic field relies on iron rich cells which respond to magnetic fields, providing qualitative (directional) and quantitative (intensity) information. Iron rich cells have been found in bacteria [Blakemore, 1975] and bees [Gould et al., 1978], and have been detected in the upper beak of pigeons, finches, robins, warbler and chickens [Falkenberg et al., 2010, Fleissner et al., 2003]. Iron-mineral rich cells, localized in the sensory dendrites [Fleissner et al., 2003], are believed to exist in all birds.
There are two proposed theories about how birds sense the geomagnetic field using iron-based magneto-reception. The first one suggests that iron-based magneto-reception depends only on magnetite (). Magnetite clusters, according to the orientation of the external magnetic field, will attract or repel each other, deforming the dendrite membrane and possibly opening or closing ion channels. On the other hand, this theory has been proposed fifteen years ago, before the discovery of maghemite () platelets in the upper beak of birds.


The second proposed theory suggests that iron-based magneto-reception depends both on magnetite and maghemite. With X-ray and histology methodologies magnetite and maghemite have been detected in the upper beak of pigeons and have been both shown to be necessary for magneto-detection [Fleissner et al., 2007]. Magnetite forms micro-clusters that are attached to the cell membrane, while maghemite crystals are arranged in chains inside the dendrites, as shown in Figure [fig:magnetite-maghemite]. It is believed that maghemite becomes magnetized thereby enhancing the magnetic field of a cell. The magnetite cluster will then experience an attractive (or repulsive) force inducing their displacement and hence the opening of ion channels.
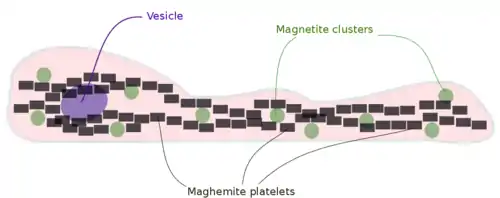
Histology has revealed that both magnetite and maghemite are present within the dendrites of the trigeminal nerve, especially in the branch that transmits sensory inputs form the upper beak to the brain. Besides this finding, researchers have shown the existence of three dendrite fields, each being responsible for coding a specific 3D orientation [Fleissner et al., 2007]. It is hypothesized that the magnetization of magnetite and maghemite caused by the geomagnetic field leads to the opening of ion channels. The information derived from magnetic fields is encoded into action potentials that reach the brain to be correctly interpreted.
There are many behavioral experiments that support this first hypothesis of avian magneto-detection. [Heyers et al., 2010] showed that changes in the magnetic field activate neurons in the trigeminal brainstem complex and that the trigeminal nerve is necessary for magneto-perception. They showed that ablation of the trigeminal nerve or removal of an external magnetic field led to reduced neuronal activation in PrV and SpV, two brain areas receiving primary inputs from the trigeminal nerve. In accordance with this study, disruption of trigeminal nerve or the attachment of a magnet to the upper beak area led to impairment of pigeons orientation [Mora et al., 2004]. Altogether, these findings strongly suggest a strict relation between the trigeminal nerve and the magnetic sensory system. However, more detailed findings are still needed.
The validity of the hypothesis here described, on how birds sense the Earth's magnetic field, has been debated by the recent finding that the believed-to-be iron-mineral structures in the trigeminal branch are in truth immune system cells called macrophages [Treiber et al., 2012]. Attempts by [Treiber et al., 2012] in finding supporting approaches to replicate electro-physiological data showing the presence of magnetite and maghemite in dendrites have failed.
The existence of iron-rich neurons in birds upper beak remains controversial. Nevertheless, the iron-based magnetic theory has not been discarded yet as many behavioral experiments, as the above cited, strongly suggest the involvement of the trigeminal nerve in magneto-perception.
Light-dependent radical-pair magneto-reception
A second hypothesis, also popular in the research field, aims at demonstrating how bird' magnetic orientation system is light dependent. This theory is supported by experiments in which avians' magnetic orientation showed an interesting dependance on a strict range of wavelengths [Wiltschko et al., 2010] . During other experiments in a cage, the utilization of full spectrum light led to birds disorientation [2]. The first research question arising from these experimental results is the origin of a structure in the birds' eye which is able to detect the geomagnetic field. How can visual and magnetic cues be separately processed is also a research topic.
The hypothesis on light-dependent magnetic sensory system states that the direction of a magnetic field is sensed by radical pair forming after photon absorption in photopigments located in the retina. Cryptochrome, a flavoprotein sensitive to blue light, has been suggested as the primary magneto-receptor in birds. Proving the validity of this theory, several cryptochrome family members have been found to be expressed in the retina of migrating birds. Moreover, their activity is highest during migratory behavior.
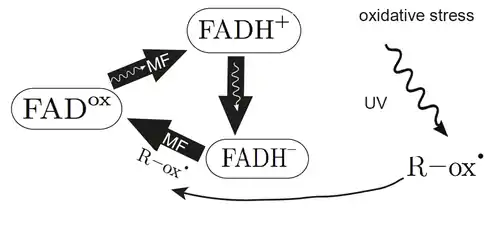
Light absorption leads to changes in the oxidation state in the cryptochrome pigment flavin adenine dinucleotide (FAD), creating an intermediate state in which the pigment, together with its electron transfer partner (tryptophan), form a radical pair. The electron spin of both radicals makes them sensitive to external magnetic fields. The different states of FAD oxidation are illustrated in the attached Figure. The homeostasis of FAD is extremely important as depending on its reduction state FAD activates different downstreaming signals.
Other important consequences and research questions arising from this hypothesis are how information reaches the brain from the retina and where light-dependent magnetic information are processed in the bird's central nervous system. Ganglion cells are the only ones transmitting information between eyes and brain, therefore magnetic information must pass through them, independently of where in the eye the magnetic cells are active.
Magnetic information gathered in the retina is then transmitted via the thalamus to a forebrain region known as Cluster N, essential to magnetic field processing. Lesions in the Cluster N have been found to affect magnetic compass orientation but not stars and sun compass orientation abilities . This forebrain region is active at night, suggesting that magnetic orientation is a primary nocturnal navigational tool, while during the day other structures are more prominent.
Given that light-dependent magnetic information is detected in the retina, it is central to address how these signals can be separated from normal vision. It is hypothesized that these systems, although in close proximity, are orientated in different directions. Rod and cone cells are oriented approximately perpendicular to the retina, whereas magnetic receptor signals depends on the angular dependence between light, receptor and magnetic field. The maximal signal speed occurs when the receptor is parallel to the geometric field. The reason why birds are able to separate magnetic and visual information is that any magnetic generated pattern moves with half the speed of the surrounding landscape.
It was believed that the magnetic compass senses in birds were strongly lateralized towards the right eye. However recent findings show that cryptochromes are located in both eyes [Mouritsen et al., 2004], Cluster N activation is similar in both brain hemispheres [Zapka et al., 2009] and neuronal pathways between eye and Cluster N are symmetrical [Heyers er al., 2007]. These results then suggest that no lateralization is present.
Despite our far from complete understanding of radical pairs in cryptochromes, they appear to fit the purpose as magneto-receptors, from a theoretical point of view. However, there are still several unresolved issues. First of all, it is unclear which of the four cryptochromes found in the bird's retina is involved in migration, nor it is known if magnetic fields can be detected in vitro by cryptochrome proteins from migratory birds. Lastly, the existence of other brain regions apart from Cluster N, which could be important for signal processing of magnetic information, needs further investigation.
Inner ear lagena
Findings suggest the existence of a third possible magneto-receptor in birds, located in the inner ear lagena organs. The lagena, found in fishes, amphibians, reptiles, birds and monotremes (but not other mammals) is defined as the third otolith organ. In pigeons the lagena lies at the base of the basilar papilla, the avian equivalent of the organ of Corti, with receptors oriented in a sagittal plane [Wu and Dickman, 2011]. The lagena is similar to its neighboring structures, the utricle and the saccule. All three detect changes in head tilt relative to gravity, translational motion and linear acceleration by means of hair cells deflection. Proving the importance of the lagena for magneto-reception, pigeons which had their lagena removed or small magnetic interference inserted into the inner ear showed compromised navigational abilities [Harada, 2002].
It is believed that detection of the geomagnetic field in the lagena, as in the trigeminal nerve, also relies on ferrimagnetic compounds [Harada et al., 2001]. Hair cells are being speculated to contain iron rich cells that senses changes in the geomagnetic field. According to this speculation a recent study [Lauwers et al., 2013] has detected iron-rich structures in both type I and type II cells in the lagena. This in turn suggests that this iron-rich particles, under the influence of the geomagnetic field, can modify the transduction of input stimuli to the brain through the deflection of hair cells leading to opening or closing of ion channels.
Despite the finding of possible magneto-sensory cells in the inner lagena, the neural pathway activated during magneto-reception is still unknown. A study has been carried out with the c-Fos transcription factor, a marker used to highlight activated neurons along the pattern of activation generated by a magnetic field. As expected, activation was detected in brain areas known to be involved in orientation and spatial memory and navigation function. Supporting the theory here discussed, much of these brain regions received information from the lagena receptor organs, while ablation of lagena led to reduced number of active neurons in those regions [Wu and Dickman, 2011].
Research issues with magnetic systems
The difficulties in identifying a magneto-receptive organ contribute to the delay in understanding how the magnetic system developed in birds and, consequently, very few is known about the molecular and genetic factors that determine this kind of sensory system.
Progress in understanding the magnetic sense has been hampered by:
- The availability of only a small number of techniques that are adequate for analyzing animal behavioral response to magnetic fields. For instance, many studies are performed in bound anesthetized animals, in which the influence of anesthesia on perception is still debated.
- The difficulty in achieving reproducible results. After the finding of
- iron-rich cells in the upper beak of birds, many electrophysiological data were
- replicated but led to different results questioning the validity of the proposed theory;
- The difficulty in implementing and carrying out new theories that might be more powerful than the ones used today. The human difficulty to understand the avian magnetic perception impedes the development of new and more efficient methods to study avian geomagnetic perception.
Redundancy of the system
It is clearly accepted that birds' orientation relays on the geomagnetic field. Still, as we have seen, no unequivocally magnetic sensitive structure nor a valid explanation to how the brain receives and interpret magnetic field information has yet been found. All the three above discussed hypotheses are plausible and well supported by many behavioral experiments. However, for all of them there are still many open questions and contradictory findings.
Taking into account all the presented evidence, it is difficult to escape the conclusion that birds' magnetoperception does not rely on a single sensory receptor but that it profits from the integration of different ones, presumably one out of the others being more prominent in specific situations.
It is believed that the iron-based magneto-reception provides quantitative or polarity data, such as the intensity of the magnetic field, whereas the cryptochrome receptors detect directional information regarding the Earth's magnetic field. It has also been shown that the iron-based magneto-reception can also control directional behavior when the radical pair process is disrupted [Wiltschko et al., 2010].
Confirming this theory, a study showed that, under blue/green light, birds oriented using cryptochrome-based detection, whereas when green/yellow light was used birds used mainly magnetite based perception [Wiltschko et al., 2012]. Hence, the idea of a possible redundancy in magnetic sensory perception can be proposed, raising a new question on how these two sources of information might be integrated as a single one.
Additionally to the sensory redundancy, studies have also shown that birds orientation does not rely exclusively on the geomagnetic field but also depend on multiple external cues. A hierarchy determining the usage of different orientation strategies has been speculated and showed that the sun or star maps are prioritized over magneto-perception. However, it is believed that information from each system is processed and integrated giving a more detailed, complex and precise representation of the environment. The way none of the elements of this redundancy can be easily shown to be prominent with regard to the other ones shows how magneto-perception can be better explained on an evolutionary basis. Redundant systems are less suitable to outside interference and can overcome situations where one important factor cannot be used or is not fully available. The convergence of multi-sensory cues helps the brain determine precise direction and location information.
References
- 1 2 Wiltschko, W. and Wiltschko, R. (2005). "Magnetic orientation and magentoreception in birds and other animals.". J Comp Physiol A 191: 675-693.
- 1 2 Wiltschko, W. and Wiltschko, R. (1995). "Migratory orientation of european robins is affected by the wavelength of light as well as by a magnetic pulse.". J Comp Physiol A, Neuroethol. Sens. Neural. Behav. Physiol 177: 363-369.
- ↑ Wiltschko, W. and Wiltschko, R. (1972). "Magnetic compass of european robins.". Science 176 (4030): 62-64.
- ↑ Viguier, C. (1882). "Le sens de l’orientation et ses organes chez les animaux et chez l’homme.". Revue Philosophique de la France et de l’Etranger: 1-36.
- ↑ Keeton, W. T.; Larkin, T. S., and Windsor, D. M. (1974). "Normal fluctuations in the earth’s magnetic field influence pigeon orientation.". Journal of comparative physiology 95(2): 95-103.
- R Blakemore. Magnetotactic bacteria. Science, 190(4212):377-9, Oct 1975.
- Gerald Falkenberg, Gerta Fleissner, Kirsten Schuchardt, Markus Kuehbacher, Peter Thalau, Henrik Mouritsen, Dominik Heyers, Gerd Wellenreuther, and Guenther Fleissner. Avian magnetoreception: elaborate iron mineral containing dendrites in the upper beak seem to be a common feature of birds. PLoS One, 5(2):e9231, 2010. doi: 10.1371/journal.pone.0009231.
- Gerta Fleissner, Elke Holtkamp-Rötzler, Marianne Hanzlik, Michael Winklhofer, Günther Fleissner, Nikolai Petersen, andWolfgang Wiltschko. Ultrastructural analysis of a putative magnetoreceptor in the beak of homing pigeons. J Comp Neurol, 458(4):350-60, Apr 2003. doi: 10.1002/cne.10579.
- Gerta Fleissner, Branko Stahl, Peter Thalau, Gerald Falkenberg, and Günther Fleissner. A novel concept of fe-mineral-based magnetoreception: histological and physicochemical data from the upper beak of homing pigeons. Naturwissenschaften, 94(8):631-42, Aug 2007. doi: 10.1007/ s00114-007-0236-0.
- Thord Fransson, Sven Jakobsson, Patrik Johansson, Cecilia Kullberg, Johan Lind, and Adrian Vallin. Bird migration: magnetic cues trigger extensive refuelling. Nature, 414(6859):35–36, 2001.
- J L Gould, J L Kirschvink, and K S Deffeyes. Bees have magnetic remanence. Science, 201(4360): 1026–8, Sep 1978. doi: 10.1126/science.201.4360.1026.
- Y Harada, M Taniguchi, H Namatame, and A Iida. Magnetic materials in otoliths of bird and fish lagena and their function. Acta Otolaryngol, 121(5):590-5, Jul 2001.
- Yasuo Harada. Experimental analysis of behavior of homing pigeons as a result of functional disorders of their lagena. Acta Otolaryngol, 122(2):132-7, Mar 2002. Dominik Heyers, Martina Manns, Harald Luksch, Onur Güntürkün, and Henrik Mouritsen. A visual pathway links brain structures active during magnetic compass orientation in migratory birds. PLoS One, 2(9):e937, 2007. doi: 10.1371/journal.pone.0000937.
- Dominik Heyers, Manuela Zapka, Mara Hoffmeister, John Martin Wild, and Henrik Mouritsen. Magnetic field changes activate the trigeminal brainstem complex in a migratory bird. Proc Natl Acad Sci U S A, 107(20):9394-9, May 2010. doi: 10.1073/pnas.0907068107.
- Mattias Lauwers, Paul Pichler, Nathaniel Bernard Edelman, Guenter Paul Resch, Lyubov Ushakova, Marion Claudia Salzer, Dominik Heyers, Martin Saunders, Jeremy Shaw, and David Anthony Keays. An iron-rich organelle in the cuticular plate of avian hair cells. Curr Biol, 23(10):924-9, May 2013. doi: 10.1016/j.cub.2013.04.025.
- Cordula V Mora, Michael Davison, J Martin Wild, and Michael M Walker. Magnetoreception and its trigeminal mediation in the homing pigeon. Nature, 432(7016):508-11, Nov 2004. doi: 10.1038/nature03077.
- Henrik Mouritsen, Ulrike Janssen-Bienhold, Miriam Liedvogel, Gesa Feenders, Julia Stalleicken, Petra Dirks, and Reto Weiler. Cryptochromes and neuronal-activity markers colocalize in the retina of migratory birds during magnetic orientation. Proc Natl Acad Sci U S A, 101(39): 14294-9, Sep 2004. doi: 10.1073/pnas.0405968101.
- Paul O’Neill. Magnetoreception and baroreception in birds. Development, growth & differentiation, 55(1):188-197, 2013.
- Thorsten Ritz, Margaret Ahmad, Henrik Mouritsen, Roswitha Wiltschko, and Wolfgang Wiltschko. Photoreceptor-based magnetoreception: optimal design of receptor molecules, cells, and neuronal processing. Journal of the Royal Society, Interface / the Royal Society, 7 Suppl 2:S135-S146, Apr 2010. ISSN 1742-5662. doi: 10.1098/rsif.2009.0456.focus. URL http://www.ncbi.nlm.nih. gov/pubmed/20129953.
- Christoph Daniel Treiber, Marion Claudia Salzer, Johannes Riegler, Nathaniel Edelman, Cristina Sugar, Martin Breuss, Paul Pichler, Herve Cadiou, Martin Saunders, Mark Lythgoe, Jeremy Shaw, and David Anthony Keays. Clusters of iron-rich cells in the upper beak of pigeons are macrophages not magnetosensitive neurons. Nature, 484(7394):367-70, Apr 2012. doi: 10.1038/ nature11046.
- Roswitha Wiltschko, Katrin Stapput, Peter Thalau, and Wolfgang Wiltschko. Directional orientation of birds by the magnetic field under different light conditions. J R Soc Interface, 7 Suppl 2: S163-77, Apr 2010. doi: 10.1098/rsif.2009.0367.focus.
- Roswitha Wiltschko, Lars Dehe, Dennis Gehring, Peter Thalau, and Wolfgang Wiltschko. Interactions between the visual and the magnetoreception system: Different effects of bichromatic light regimes on the directional behavior of migratory birds. J Physiol Paris, Apr 2012. doi: 10.1016/j.jphysparis.2012.03.003.
- Le-Qing Wu and J David Dickman. Magnetoreception in an avian brain in part mediated by inner ear lagena. Curr Biol, 21(5):418-23, Mar 2011. doi: 10.1016/j.cub.2011.01.058.
- Manuela Zapka, Dominik Heyers, Christine M Hein, Svenja Engels, Nils-Lasse Schneider, Jörg Hans, Simon Weiler, David Dreyer, Dmitry Kishkinev, J Martin Wild, and Henrik Mouritsen. Visual but not trigeminal mediation of magnetic compass information in a migratory bird. Nature, 461 (7268):1274-7, Oct 2009. doi: 10.1038/nature08528.