TCF7L2
Transcription factor 7-like 2 (T-cell specific, HMG-box), also known as TCF7L2 or TCF4, is a protein acting as a transcription factor that, in humans, is encoded by the TCF7L2 gene.[5][6] The TCF7L2 gene is located on chromosome 10q25.2–q25.3, contains 19 exons.[7][8] As a member of the TCF family, TCF7L2 can form a bipartite transcription factor and influence several biological pathways, including the Wnt signalling pathway.[9]
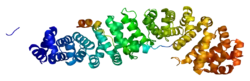




Single-nucleotide polymorphisms (SNPs) in this gene are especially known to be linked to higher risk to develop type 2 diabetes,[9] gestational diabetes,[10] multiple neurodevelopmental disorders[11][12] including schizophrenia[13][14] and autism spectrum disorder,[15][16] as well as other diseases.[17][18] The SNP rs7903146, within the TCF7L2 gene, is, to date, the most significant genetic marker associated with type 2 diabetes risk.[19]
Function
TCF7L2 is a transcription factor influencing the transcription of several genes thereby exerting a large variety of functions within the cell. It is a member of the TCF family that can form a bipartite transcription factor (β-catenin/TCF) alongside β-catenin.[9] Bipartite transcription factors can have large effects on the Wnt signalling pathway.[9] Stimulation of the Wnt signaling pathway leads to the association of β-catenin with BCL9, translocation to the nucleus, and association with TCF7L2,[21] which in turn results in the activation of Wnt target genes. The activation of the Wnt target genes specifically represses proglucagon synthesis in enteroendocrine cells.[9][8] The repression of TCF7L2 using HMG-box repressor (HBP1) inhibits Wnt signalling.[9] Therefore, TCF7L2 is an effector in the Wnt signalling pathway. TCF7L2's role in glucose metabolism is expressed in many tissues such as gut, brain, liver, and skeletal muscle. However, TCF7L2 does not directly regulate glucose metabolism in β-cells, but regulates glucose metabolism in pancreatic and liver tissues.[22] That said, TCF7L2 directly regulates the expression of multiple transcription factors, axon guidance cues, cell adhesion molecules and ion channels in the thalamus.[23]
The TCF7L2 gene encoding the TCF7L2 transcription factor, exhibits multiple functions through its polymorphisms and thus, is known as a pleiotropic gene. Type 2 diabetes T2DM susceptibility is exhibited in carriers of TCF7L2 rs7903146C>T[24][25] and rs290481T>C[25] polymorphisms.[24][25] TCF7L2 rs290481T>C polymorphism, however, has shown no significant correlation to the susceptibility to gestational diabetes mellitus (GDM) in a Chinese Han population, whereas the T alleles of rs7903146[25] and rs1799884[10] increase susceptibility to GDM in the Chinese Han population.[25][10] The difference in effects of the different polymorphisms of the gene indicate that the gene is indeed pleiotropic.
Structure
The TCF7L2 gene, encoding the TCF7L2 protein, is located on chromosome 10q25.2-q25.3. The gene contains 19 exons.[7][8] Of the 19 exons, 5 are alternative.[8] The TCF7L2 protein contains 619 amino acids and its molecular mass is 67919 Da.[26] TCF7L2's secondary structure is a helix-turn-helix structure.[27]
Tissue distribution
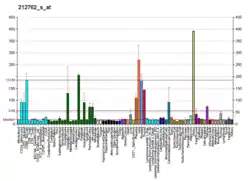
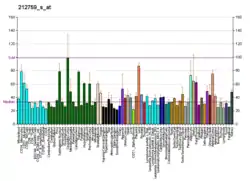
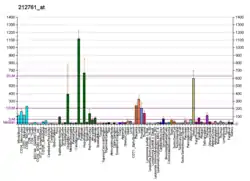
TCF7L2 is primarily expressed in brain (mainly in the diencephalon, including especially high in the thalamus[23][28][29]), liver, intestine and fat cells. It does not primarily operate in the β-cells in the pancreas.[30]
Clinical significance
Type 2 Diabetes
Several single nucleotide polymorphisms within the TCF7L2 gene have been associated with type 2 diabetes. Studies conducted by Ravindranath Duggirala and Michael Stern at The University of Texas Health Science Center at San Antonio were the first to identify strong linkage for type 2 diabetes at a region on Chromosome 10 in Mexican Americans [31] This signal was later refined by Struan Grant and colleagues at DeCODE genetics and isolated to the TCF7L2 gene.[32] The molecular and physiological mechanisms underlying the association of TCF7L2 with type 2 diabetes are under active investigation, but it is likely that TCF7L2 has important biological roles in multiple metabolic tissues, including the pancreas, liver and adipose tissue.[30][33] TCF7L2 polymorphisms can increase susceptibility to type 2 diabetes by decreasing the production of glucagon-like peptide-1 (GLP-1).[9]
Gestational Diabetes (GDM)
TCF7L2 modulates pancreatic islet β-cell function strongly implicating its significant association with GDM risk.[10] T alleles of rs7903146[25] and rs1799884[10] TCF7L2 polymorphisms increase susceptibility to GDM in the Chinese Han population.[25][10]
Cancer
TCF7L2 plays a role in colorectal cancer.[17] A frameshift mutation of TCF7L2 provided evidence that TCF7L2 is implicated in colorectal cancer.[34][35] The silencing of TCF7L2 in KM12 colorectal cancer cells provided evidence that TCF7L2 played a role in proliferation and metastasis of cancer cells in colorectal cancer.[17]
Variants of the gene are most likely involved in many other cancer types.[36] TCF7L2 is indirectly involved in prostate cancer through its role in activating the PI3K/Akt pathway, a pathway involved in prostate cancer.[37]
Neurodevelopmental disorders
Single nucleotide polymorphisms (SNPs) in TCF7L2 gene have shown an increase in susceptibility to schizophrenia in Arab, European and Chinese Han populations. In the Chinese Han population, SNP rs12573128[14] in TCF7L2 is the variant that was associated with an increase in schizophrenia risk. This marker is used as a pre-diagnostic marker for schizophrenia.[14] TCF7L2 has also been reported as a risk gene in autism spectrum disorder[38] and has been linked to it in recent large-scale genetic studies.[15][16]
The mechanism behind TCF7L2's involvement in the emergence of neurodevelopmental disorders is not fully understood, as there have been few studies characterizing its role in brain development in detail. It was shown that during embryogenesis TCF7L2 is involved in the development of fish-specific habenula asymmetry in Danio rerio,[39][40] and that the dominant negative TCF7L2 isoform influences cephalic separation in the embryo by inhibiting the posteriorizing effect of the Wnt pathway.[41] It was also shown that in Tcf7l2 knockout mice the number of proliferating cells in cortical neural progenitor cells is reduced.[42] In contrast, no such effect was found in the midbrain.[43]
More recently it was shown that TCF7L2 plays a crucial role in both the embryonic development and postnatal maturation of the thalamus through direct and indirect regulation of many genes previously reported to be important for both processes.[23] In late gestation TCF7L2 regulates the expression of many thalamus-enriched transcription factors (e.g. Foxp2, Rora, Mef2a, Lef1, Prox1), axon guidance molecules (e.g. Epha1, Epha4, Ntng1, Epha8) and cell adhesion molecules (e.g. Cdh6, Cdh8, Cdhr1). Accordingly, a total knockout of Tcf7l2 in mice leads to improper growth of thalamocortical axons, changed anatomy and improper sorting of the cells in the thalamo-habenular region.[23] In the early postnaral period TCF7L2 starts to regulate the expression of many genes necessary for the acquisition of characteristic excitability patterns in the thalamus, mainly ion channels, neurotransmitters and their receptors and synaptic vescicle proteins (e.g. Cacna1g, Kcnc2, Slc17a7, Grin2b), and an early postnatal knockout of Tcf7l2 in mouse thalamus leads to significant reduction in the number and frequency of action potentials generated by the thalamocortical neurons.[23] The mechanism that leads to the change in TCF7L2 target genes between gestation and early postnatal period is unknown. It is likely that a perinatal change in the proportion of TCF7L2 isoforms expressed in the thalamus is partially responsible.[28] Abnormalities in the anatomy of the thalamus and the activity of its connections to the cerebral cortex are frequently detected in patients with schizophrenia [44][45][46][47] and autism.[48][49][50][51] Such abnormalities could arise from developmental aberrations in patients with unfavouralbe mutations of TCF7L2, further strengthening the link between TCF7L2 and neurodevelopmental disorders.
Multiple sclerosis
TCF7L2 is downstream of the WNT/β-catenin pathways. The activation of the WNT/β-catenin pathways have been associated demyelination in multiple sclerosis.[18] TCF7L2 is unregulated during early remyelination, leading scientists to believe that it is involved in remyelination.[18] TCF7L2 could act in dependence or independent of the WNT/β-catenin pathways.[18]
Model organisms
Model organisms have been used in the study of TCF7L2 function. A conditional knockout mouse line called Tcf7l2tm1a(EUCOMM)Wtsi was generated at the Wellcome Trust Sanger Institute.[52] Male and female animals underwent a standardized phenotypic screen[53] to determine the effects of deletion.[54][55][56][57] Additional screens performed: - In-depth immunological phenotyping[58]
Variations of the protein encoding gene are found in rats, zebra fish, drosophila, and budding yeast.[59] Therefore, all of those organisms can be used as model organisms in the study of TCF7L2 function.
Nomenclature
TCF7L2 is the symbol officially approved by the HUGO Gene Nomenclature Committee for the transcription factor 4 gene (TCF4).
See also
References
- GRCh38: Ensembl release 89: ENSG00000148737 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000024985 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Entrez Gene: TCF7L2".
- Castrop J, van Norren K, Clevers H (February 1992). "A gene family of HMG-box transcription factors with homology to TCF-1". Nucleic Acids Research. 20 (3): 611. doi:10.1093/nar/20.3.611. PMC 310434. PMID 1741298.
- "TCF7L2 transcription factor 7 like 2 [Homo sapiens (human)] - Gene - NCBI". www.ncbi.nlm.nih.gov. Retrieved 2017-11-30.
- Online Mendelian Inheritance in Man (OMIM): TRANSCRIPTION FACTOR 7-LIKE 2;TCF7L2 - 602228{
- Jin T, Liu L (November 2008). "The Wnt signaling pathway effector TCF7L2 and type 2 diabetes mellitus". Molecular Endocrinology. 22 (11): 2383–2392. doi:10.1210/me.2008-0135. PMID 18599616.
- Zhang C, Bao W, Rong Y, Yang H, Bowers K, Yeung E, Kiely M (2013-05-19). "Genetic variants and the risk of gestational diabetes mellitus: a systematic review". Human Reproduction Update. 19 (4): 376–390. doi:10.1093/humupd/dmt013. PMC 3682671. PMID 23690305.
- Deciphering Developmental Disorders Study; et al. (March 2015). "Large-scale discovery of novel genetic causes of developmental disorders". Nature. 519 (7542): 223–228. Bibcode:2015Natur.519..223T. doi:10.1038/nature14135. PMC 5955210. PMID 25533962.
- Deciphering Developmental Disorders Study; et al. (February 2017). "Prevalence and architecture of de novo mutations in developmental disorders". Nature. 542 (7642): 433–438. Bibcode:2017Natur.542..433M. doi:10.1038/nature21062. PMC 6016744. PMID 28135719.
- Hansen T, Ingason A, Djurovic S, Melle I, Fenger M, Gustafsson O, et al. (July 2011). "At-risk variant in TCF7L2 for type II diabetes increases risk of schizophrenia". Biological Psychiatry. 70 (1): 59–63. doi:10.1016/j.biopsych.2011.01.031. PMID 21414605. S2CID 42205809.
- Liu L, Li J, Yan M, Li J, Chen J, Zhang Y, et al. (April 2017). "TCF7L2 polymorphisms and the risk of schizophrenia in the Chinese Han population". Oncotarget. 8 (17): 28614–28620. doi:10.18632/oncotarget.15603. PMC 5438676. PMID 28404897.
- Wang T, Hoekzema K, Vecchio D, Wu H, Sulovari A, Coe BP, et al. (October 2020). "Large-scale targeted sequencing identifies risk genes for neurodevelopmental disorders". Nature Communications. 11 (1): 4932. doi:10.1038/s41467-020-18723-y. PMC 7530681. PMID 33004838.
- Satterstrom FK, Kosmicki JA, Wang J, Breen MS, De Rubeis S, An JY, et al. (February 2020). "Large-Scale Exome Sequencing Study Implicates Both Developmental and Functional Changes in the Neurobiology of Autism". Cell. 180 (3): 568–584.e23. doi:10.1016/j.cell.2019.12.036. PMC 7250485. PMID 31981491.
- Torres S, García-Palmero I, Marín-Vicente C, Bartolomé RA, Calviño E, Fernández-Aceñero MJ, Casal JI (January 2018). "Proteomic Characterization of Transcription and Splicing Factors Associated with a Metastatic Phenotype in Colorectal Cancer". Journal of Proteome Research. 17 (1): 252–264. doi:10.1021/acs.jproteome.7b00548. hdl:10261/160082. PMID 29131639.
- Vallée A, Vallée JN, Guillevin R, Lecarpentier Y (May 2018). "Interactions Between the Canonical WNT/Beta-Catenin Pathway and PPAR Gamma on Neuroinflammation, Demyelination, and Remyelination in Multiple Sclerosis". Cellular and Molecular Neurobiology. 38 (4): 783–795. doi:10.1007/s10571-017-0550-9. PMID 28905149. S2CID 4620853.
- Vaquero AR, Ferreira NE, Omae SV, Rodrigues MV, Teixeira SK, Krieger JE, Pereira AC (October 2012). "Using gene-network landscape to dissect genotype effects of TCF7L2 genetic variant on diabetes and cardiovascular risk". Physiological Genomics. 44 (19): 903–914. doi:10.1152/physiolgenomics.00030.2012. PMID 22872755. S2CID 35065699.
- PDB: 2GL7; Sampietro J, Dahlberg CL, Cho US, Hinds TR, Kimelman D, Xu W (October 2006). "Crystal structure of a beta-catenin/BCL9/Tcf4 complex". Molecular Cell. 24 (2): 293–300. doi:10.1016/j.molcel.2006.09.001. PMID 17052462.
- Lee JM, Dedhar S, Kalluri R, Thompson EW (March 2006). "The epithelial-mesenchymal transition: new insights in signaling, development, and disease". The Journal of Cell Biology. 172 (7): 973–981. doi:10.1083/jcb.200601018. PMC 2063755. PMID 16567498.
- Facchinello N, Tarifeño-Saldivia E, Grisan E, Schiavone M, Peron M, Mongera A, et al. (August 2017). "Tcf7l2 plays pleiotropic roles in the control of glucose homeostasis, pancreas morphology, vascularization and regeneration". Scientific Reports. 7 (1): 9605. Bibcode:2017NatSR...7.9605F. doi:10.1038/s41598-017-09867-x. PMC 5575064. PMID 28851992.
- Lipiec MA, Bem J, Koziński K, Chakraborty C, Urban-Ciećko J, Zajkowski T, et al. (August 2020). "TCF7L2 regulates postmitotic differentiation programmes and excitability patterns in the thalamus". Development. 147 (16): dev.190181. doi:10.1242/dev.190181. PMC 7473649. PMID 32675279.
- Chen Y, Zhao Y, Li YB, Wang YJ, Zhang GZ (January 2018). "Detection of SNPs of T2DM susceptibility genes by a ligase detection reaction-fluorescent nanosphere technique". Analytical Biochemistry. 540–541 (Supplement C): 38–44. doi:10.1016/j.ab.2017.11.003. PMID 29128291.
- Zhu L, Xie Z, Lu J, Hao Q, Kang M, Chen S, et al. (September 2017). "TCF7L2 rs290481 T>C polymorphism is associated with an increased risk of type 2 diabetes mellitus and fasting plasma glucose level". Oncotarget. 8 (44): 77000–77008. doi:10.18632/oncotarget.20300. PMC 5652758. PMID 29100364.
- Database, GeneCards Human Gene. "TCF7L2 Gene - GeneCards | TF7L2 Protein | TF7L2 Antibody". www.genecards.org. Retrieved 2017-11-30.
- "TCF7L2 - Transcription factor 7-like 2 - Homo sapiens (Human) - TCF7L2 gene & protein". www.uniprot.org. Retrieved 2017-11-30.
- Nagalski A, Irimia M, Szewczyk L, Ferran JL, Misztal K, Kuznicki J, Wisniewska MB (November 2013). "Postnatal isoform switch and protein localization of LEF1 and TCF7L2 transcription factors in cortical, thalamic, and mesencephalic regions of the adult mouse brain". Brain Structure & Function. 218 (6): 1531–1549. doi:10.1007/s00429-012-0474-6. PMC 3825142. PMID 23152144.
- Murray KD, Rubin CM, Jones EG, Chalupa LM (November 2008). "Molecular correlates of laminar differences in the macaque dorsal lateral geniculate nucleus". The Journal of Neuroscience. 28 (46): 12010–12022. doi:10.1523/JNEUROSCI.3800-08.2008. PMC 2613947. PMID 19005066.
- Nobrega MA (March 2013). "TCF7L2 and glucose metabolism: time to look beyond the pancreas". Diabetes. 62 (3): 706–708. doi:10.2337/db12-1418. PMC 3581232. PMID 23431017.
- Duggirala R, Blangero J, Almasy L, Dyer TD, Williams KL, Leach RJ, et al. (April 1999). "Linkage of type 2 diabetes mellitus and of age at onset to a genetic location on chromosome 10q in Mexican Americans". American Journal of Human Genetics. 64 (4): 1127–1140. doi:10.1086/302316. PMC 1377837. PMID 10090898.
- Grant SF, Thorleifsson G, Reynisdottir I, Benediktsson R, Manolescu A, Sainz J, et al. (March 2006). "Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes". Nature Genetics. 38 (3): 320–323. doi:10.1038/ng1732. PMID 16415884. S2CID 28825825.
- Jin T (June 2016). "Current Understanding on Role of the Wnt Signaling Pathway Effector TCF7L2 in Glucose Homeostasis". Endocrine Reviews. 37 (3): 254–277. doi:10.1210/er.2015-1146. PMID 27159876.
- Slattery ML, Folsom AR, Wolff R, Herrick J, Caan BJ, Potter JD (April 2008). "Transcription factor 7-like 2 polymorphism and colon cancer". Cancer Epidemiology, Biomarkers & Prevention. 17 (4): 978–982. doi:10.1158/1055-9965.EPI-07-2687. PMC 2587179. PMID 18398040.
- Hazra A, Fuchs CS, Chan AT, Giovannucci EL, Hunter DJ (November 2008). "Association of the TCF7L2 polymorphism with colorectal cancer and adenoma risk". Cancer Causes & Control. 19 (9): 975–980. doi:10.1007/s10552-008-9164-3. PMC 2719293. PMID 18478343.
- Tang W, Dodge M, Gundapaneni D, Michnoff C, Roth M, Lum L (July 2008). "A genome-wide RNAi screen for Wnt/beta-catenin pathway components identifies unexpected roles for TCF transcription factors in cancer". Proceedings of the National Academy of Sciences of the United States of America. 105 (28): 9697–9702. Bibcode:2008PNAS..105.9697T. doi:10.1073/pnas.0804709105. PMC 2453074. PMID 18621708.
- Sun P, Xiong H, Kim TH, Ren B, Zhang Z (February 2006). "Positive inter-regulation between beta-catenin/T cell factor-4 signaling and endothelin-1 signaling potentiates proliferation and survival of prostate cancer cells". Molecular Pharmacology. 69 (2): 520–531. doi:10.1124/mol.105.019620. PMID 16291872. S2CID 10148857.
- "SFARI Gene Databse - TCF7L2". Retrieved 2022-07-16.
- Beretta CA, Dross N, Bankhead P, Carl M (September 2013). "The ventral habenulae of zebrafish develop in prosomere 2 dependent on Tcf7l2 function". Neural Development. 8 (1): 19. doi:10.1186/1749-8104-8-19. PMC 3827927. PMID 24067090.
- Hüsken U, Stickney HL, Gestri G, Bianco IH, Faro A, Young RM, et al. (October 2014). "Tcf7l2 is required for left-right asymmetric differentiation of habenular neurons". Current Biology. 24 (19): 2217–2227. doi:10.1016/j.cub.2014.08.006. PMC 4194317. PMID 25201686.
- Vacik T, Stubbs JL, Lemke G (September 2011). "A novel mechanism for the transcriptional regulation of Wnt signaling in development". Genes & Development. 25 (17): 1783–1795. doi:10.1101/gad.17227011. PMC 3175715. PMID 21856776.
- Chodelkova O, Masek J, Korinek V, Kozmik Z, Machon O (May 2018). "Tcf7L2 is essential for neurogenesis in the developing mouse neocortex". Neural Development. 13 (1): 8. doi:10.1186/s13064-018-0107-8. PMC 5946422. PMID 29751817.
- Lee M, Yoon J, Song H, Lee B, Lam DT, Yoon J, et al. (April 2017). "Tcf7l2 plays crucial roles in forebrain development through regulation of thalamic and habenular neuron identity and connectivity". Developmental Biology. 424 (1): 62–76. doi:10.1016/j.ydbio.2017.02.010. PMID 28219675.
- de Zwarte SM, Brouwer RM, Tsouli A, Cahn W, Hillegers MH, Hulshoff Pol HE, et al. (October 2019). "Running in the Family? Structural Brain Abnormalities and IQ in Offspring, Siblings, Parents, and Co-twins of Patients with Schizophrenia". Schizophrenia Bulletin. 45 (6): 1209–1217. doi:10.1093/schbul/sby182. PMC 6811835. PMID 30597053.
- Giraldo-Chica M, Woodward ND (February 2017). "Review of thalamocortical resting-state fMRI studies in schizophrenia". Schizophrenia Research. Pathologies of the Thalamus in Schizophrenia. 180: 58–63. doi:10.1016/j.schres.2016.08.005. PMC 5297399. PMID 27531067.
- Hua J, Blair NI, Paez A, Choe A, Barber AD, Brandt A, et al. (April 2019). "Altered functional connectivity between sub-regions in the thalamus and cortex in schizophrenia patients measured by resting state BOLD fMRI at 7T". Schizophrenia Research. 206: 370–377. doi:10.1016/j.schres.2018.10.016. PMC 6500777. PMID 30409697.
- van Erp TG, Walton E, Hibar DP, Schmaal L, Jiang W, Glahn DC, et al. (November 2018). "Cortical Brain Abnormalities in 4474 Individuals With Schizophrenia and 5098 Control Subjects via the Enhancing Neuro Imaging Genetics Through Meta Analysis (ENIGMA) Consortium". Biological Psychiatry. 84 (9): 644–654. doi:10.1016/j.biopsych.2018.04.023. PMC 6177304. PMID 29960671.
- Ayub R, Sun KL, Flores RE, Lam VT, Jo B, Saggar M, Fung LK (February 2021). "Thalamocortical connectivity is associated with autism symptoms in high-functioning adults with autism and typically developing adults". Translational Psychiatry. 11 (1): 93. doi:10.1038/s41398-021-01221-0. PMC 7859407. PMID 33536431.
- Schuetze M, Park MT, Cho IY, MacMaster FP, Chakravarty MM, Bray SL (October 2016). "Morphological Alterations in the Thalamus, Striatum, and Pallidum in Autism Spectrum Disorder". Neuropsychopharmacology. 41 (11): 2627–2637. doi:10.1038/npp.2016.64. PMC 5026732. PMID 27125303.
- Tomasi D, Volkow ND (February 2019). "Reduced Local and Increased Long-Range Functional Connectivity of the Thalamus in Autism Spectrum Disorder". Cerebral Cortex. 29 (2): 573–585. doi:10.1093/cercor/bhx340. PMC 6319176. PMID 29300843.
- Woodward ND, Giraldo-Chica M, Rogers B, Cascio CJ (January 2017). "Thalamocortical dysconnectivity in autism spectrum disorder: An analysis of the Autism Brain Imaging Data Exchange". Biological Psychiatry. Cognitive Neuroscience and Neuroimaging. 2 (1): 76–84. doi:10.1016/j.bpsc.2016.09.002. PMC 5455796. PMID 28584881.
- Gerdin AK (2010). "The Sanger Mouse Genetics Programme: high throughput characterisation of knockout mice". Acta Ophthalmologica. 88: 925–7. doi:10.1111/j.1755-3768.2010.4142.x. S2CID 85911512.
- "International Mouse Phenotyping Consortium".
- Skarnes WC, Rosen B, West AP, Koutsourakis M, Bushell W, Iyer V, et al. (June 2011). "A conditional knockout resource for the genome-wide study of mouse gene function". Nature. 474 (7351): 337–342. doi:10.1038/nature10163. PMC 3572410. PMID 21677750.
- Dolgin E (June 2011). "Mouse library set to be knockout". Nature. 474 (7351): 262–263. doi:10.1038/474262a. PMID 21677718.
- Collins FS, Rossant J, Wurst W (January 2007). "A mouse for all reasons". Cell. 128 (1): 9–13. doi:10.1016/j.cell.2006.12.018. PMID 17218247. S2CID 18872015.
- White JK, Gerdin AK, Karp NA, Ryder E, Buljan M, Bussell JN, et al. (July 2013). "Genome-wide generation and systematic phenotyping of knockout mice reveals new roles for many genes". Cell. 154 (2): 452–464. doi:10.1016/j.cell.2013.06.022. PMC 3717207. PMID 23870131.
- "Infection and Immunity Immunophenotyping (3i) Consortium".
- "MARRVEL: Search Result". marrvel.org. Retrieved 2017-11-30.
Further reading
- Segditsas S, Tomlinson I (December 2006). "Colorectal cancer and genetic alterations in the Wnt pathway". Oncogene. 25 (57): 7531–7537. doi:10.1038/sj.onc.1210059. PMID 17143297.
- Florez JC (July 2007). "The new type 2 diabetes gene TCF7L2". Current Opinion in Clinical Nutrition and Metabolic Care. 10 (4): 391–396. doi:10.1097/MCO.0b013e3281e2c9be. PMID 17563454. S2CID 21362394.
- Maruyama K, Sugano S (January 1994). "Oligo-capping: a simple method to replace the cap structure of eukaryotic mRNAs with oligoribonucleotides". Gene. 138 (1–2): 171–174. doi:10.1016/0378-1119(94)90802-8. PMID 8125298.
- Korinek V, Barker N, Morin PJ, van Wichen D, de Weger R, Kinzler KW, et al. (March 1997). "Constitutive transcriptional activation by a beta-catenin-Tcf complex in APC-/- colon carcinoma". Science. 275 (5307): 1784–1787. doi:10.1126/science.275.5307.1784. hdl:20.500.11755/27e2349d-dfbe-4458-9a4e-3fa15f0d2420. PMID 9065401. S2CID 33935423.
- Suzuki Y, Yoshitomo-Nakagawa K, Maruyama K, Suyama A, Sugano S (October 1997). "Construction and characterization of a full length-enriched and a 5'-end-enriched cDNA library". Gene. 200 (1–2): 149–156. doi:10.1016/S0378-1119(97)00411-3. PMID 9373149.
- He TC, Sparks AB, Rago C, Hermeking H, Zawel L, da Costa LT, et al. (September 1998). "Identification of c-MYC as a target of the APC pathway". Science. 281 (5382): 1509–1512. Bibcode:1998Sci...281.1509H. doi:10.1126/science.281.5382.1509. PMID 9727977.
- Barker N, Huls G, Korinek V, Clevers H (January 1999). "Restricted high level expression of Tcf-4 protein in intestinal and mammary gland epithelium". The American Journal of Pathology. 154 (1): 29–35. doi:10.1016/S0002-9440(10)65247-9. PMC 1853446. PMID 9916915.
- Omer CA, Miller PJ, Diehl RE, Kral AM (March 1999). "Identification of Tcf4 residues involved in high-affinity beta-catenin binding". Biochemical and Biophysical Research Communications. 256 (3): 584–590. doi:10.1006/bbrc.1999.0379. PMID 10080941.
- Giannini AL, Vivanco MM, Kypta RM (March 2000). "Analysis of beta-catenin aggregation and localization using GFP fusion proteins: nuclear import of alpha-catenin by the beta-catenin/Tcf complex". Experimental Cell Research. 255 (2): 207–220. doi:10.1006/excr.1999.4785. PMID 10694436.
- Duval A, Busson-Leconiat M, Berger R, Hamelin R (2000). "Assignment of the TCF-4 gene (TCF7L2) to human chromosome band 10q25.3". Cytogenetics and Cell Genetics. 88 (3–4): 264–265. doi:10.1159/000015534. PMID 10828605. S2CID 13148464.
- Duval A, Rolland S, Tubacher E, Bui H, Thomas G, Hamelin R (July 2000). "The human T-cell transcription factor-4 gene: structure, extensive characterization of alternative splicings, and mutational analysis in colorectal cancer cell lines". Cancer Research. 60 (14): 3872–3879. PMID 10919662.
- Brantjes H, Roose J, van De Wetering M, Clevers H (April 2001). "All Tcf HMG box transcription factors interact with Groucho-related co-repressors". Nucleic Acids Research. 29 (7): 1410–1419. doi:10.1093/nar/29.7.1410. PMC 31284. PMID 11266540.
- Palacino JJ, Murphy MP, Murayama O, Iwasaki K, Fujiwara M, Takashima A, et al. (October 2001). "Presenilin 1 regulates beta-catenin-mediated transcription in a glycogen synthase kinase-3-independent fashion". The Journal of Biological Chemistry. 276 (42): 38563–38569. doi:10.1074/jbc.M105376200. PMID 11504726.
- Miravet S, Piedra J, Miró F, Itarte E, García de Herreros A, Duñach M (January 2002). "The transcriptional factor Tcf-4 contains different binding sites for beta-catenin and plakoglobin". The Journal of Biological Chemistry. 277 (3): 1884–1891. doi:10.1074/jbc.M110248200. PMID 11711551.
- Graham TA, Ferkey DM, Mao F, Kimelman D, Xu W (December 2001). "Tcf4 can specifically recognize beta-catenin using alternative conformations". Nature Structural Biology. 8 (12): 1048–1052. doi:10.1038/nsb718. PMID 11713475. S2CID 33878077.
- Poy F, Lepourcelet M, Shivdasani RA, Eck MJ (December 2001). "Structure of a human Tcf4-beta-catenin complex". Nature Structural Biology. 8 (12): 1053–1057. doi:10.1038/nsb720. PMID 11713476. S2CID 24798619.
- Thiele A, Wasner M, Müller C, Engeland K, Hauschildt S (December 2001). "Regulation and possible function of beta-catenin in human monocytes". Journal of Immunology. 167 (12): 6786–6793. doi:10.4049/jimmunol.167.12.6786. PMID 11739494.
- Marchenko GN, Marchenko ND, Leng J, Strongin AY (April 2002). "Promoter characterization of the novel human matrix metalloproteinase-26 gene: regulation by the T-cell factor-4 implies specific expression of the gene in cancer cells of epithelial origin". The Biochemical Journal. 363 (Pt 2): 253–262. doi:10.1042/0264-6021:3630253. PMC 1222473. PMID 11931652.
- Leung JY, Kolligs FT, Wu R, Zhai Y, Kuick R, Hanash S, et al. (June 2002). "Activation of AXIN2 expression by beta-catenin-T cell factor. A feedback repressor pathway regulating Wnt signaling". The Journal of Biological Chemistry. 277 (24): 21657–21665. doi:10.1074/jbc.M200139200. PMID 11940574.
External links
- TCF7L2 here called TCF4 features on this Wnt pathway web site: Wnt signalling molecules TCFs
- Structure determination of TCF7L2: PDB entry 2GL7 and related publication on PubMed
- PubMed GeneRIFs (summaries of related scientific publications) -
- Weizmann Institute GeneCard for TCF7L2
- Overview of all the structural information available in the PDB for UniProt: Q9NQB0 (Transcription factor 7-like 2) at the PDBe-KB.
PDB gallery | |
|---|---|
|



