Phosphoribosylglycinamide formyltransferase
Phosphoribosylglycinamide formyltransferase (EC 2.1.2.2, 2-amino-N-ribosylacetamide 5'-phosphate transformylase, GAR formyltransferase, GAR transformylase, glycinamide ribonucleotide transformylase, GAR TFase, 5,10-methenyltetrahydrofolate:2-amino-N-ribosylacetamide ribonucleotide transformylase) is an enzyme with systematic name 10-formyltetrahydrofolate:5'-phosphoribosylglycinamide N-formyltransferase.[1][2][3] This enzyme catalyses the following chemical reaction
- 10-formyltetrahydrofolate + N1-(5-phospho-D-ribosyl)glycinamide tetrahydrofolate + N2-formyl-N1-(5-phospho-D-ribosyl)glycinamide

| Phosphoribosylglycinamide formyltransferase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
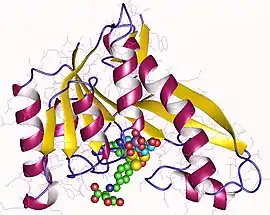 GAR formyltransferase monomer, Human | |||||||||
| Identifiers | |||||||||
| EC no. | 2.1.2.2 | ||||||||
| CAS no. | 2604945 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| |||||||||
This THF dependent enzyme catalyzes a nucleophilic acyl substitution of the formyl group from 10-formyltetrahydrofolate (fTHF) to N1-(5-phospho-D-ribosyl)glycinamide (GAR) to form N2-formyl-N1-(5-phospho-D-ribosyl)glycinamide (fGAR) as shown above.[4] This reaction plays an important role in the formation of purine through the de novo purine biosynthesis pathway. This pathway creates inosine monophosphate (IMP), a precursor to adenosine monophosphate (AMP) and guanosine monophosphate (GMP). AMP is a building block for important energy carriers such as ATP, NAD+ and FAD, and signaling molecules such as cAMP. GARTfase's role in de novo purine biosynthesis makes it a target for anti-cancer drugs[5] and its overexpression during postnatal development has been connected to Down syndrome.[6] There are two known types of genes encoding GAR transformylase in E.coli: purN and purT, while only purN is found in humans.[7] Many residues in the active site are conserved across bacterial, yeast, avian and human enzymes.[8]
Enzyme Structure
In humans, GARTfase is part of trifunctional enzyme which also includes glycinamide ribonucleotide synthase (GARS) and aminoimidazole ribonucleotide synthetase (AIRS). This protein (110kDa) catalyzes steps 2, 3 and 5 of de novo purine biosynthesis. The proximity of these enzyme units and flexibility of the protein serves to increase pathway throughput. GARTfase is located on the C-terminal end of the protein.[10]
Human GARTfase has been crystallized by vapor-diffusion sitting drop method and imaged at the Stanford Synchrotron Radiation Laboratory (SSRL) by at least two groups.[5][11]
The structure can be described by two subdomains which are connected by a seven-stranded beta sheet. The N- terminal domain consists of a Rossman type mononucleotide fold, with a four strand part of the beta sheet surrounded on each side by two alpha helices. The beta sheet continues into the C terminal domain, where on one side it is covered by a long alpha helix and on the other it is partially exposed to solvent. It is the cleft between the two subdomains where the active site lies.[8]
The cleft consists of the GAR binding site and the folate binding pocket. The folate binding pocket is delineated by pteridine-binding cleft, the formyl transfer region and the benzoylglutamate region which bind thepteridine head and a benzoylglutamate tail connected by a formyl bound nitrogen of fTHF. This folate binding region has been the subject of much research because its inhibition by small molecules has led to the discovery of antineoplastic drugs. The folate binding loop has been shown to change conformation depending on the pH of solution and as such Human GAR transformylase shows highest activity around pH 7.5-8. Lower pH (~4.2) conditions change the conformation of the substrate (GAR) binding loops as well.[11]
Mechanism
Mechanism of purN GARTfase
Klein et al first suggested a water molecule assisted mechanism. A single water molecule possibly held in place by hydrogen bonding with the carboxylate group of the persistent Asp144 residue transfers protons from the GAR-N to the THF-N. The nucleophilic nitrogen on the terminal amino group of GAR attacks the carbonyl carbon of the formyl group on THF pushing negative charge onto the oxygen. Klein suggests that His108 stabilizes the transition state by hydrogen bonding with the negatively charged oxygen and that the reformation of the carbonyl double bond results in breaking the THF-N - formyl bond. Calculations by Qiao et al suggest that the water assisted stepwise proton transfer from Gar-N to THF-N is 80-100 kj/mol more favorable than the concerted transfer suggested by Klein. The mechanism shown is suggested by Qiao et al, whom admittedly did not consider surrounding residues in their calculations.[12][13] Much of the early active site mapping on GAR TFase was determined with the bacterial enzyme owing to the quantity available from its overexpression in E. coli.[14] Using a bromoacetyl dideazafolate affinity analog James Inglese and colleagues first identified Asp144 as an active site residue likely involved in the formyl transfer mechanism.[15]
Mechanism of purT GARTfase
Studies of the purT variant of GAR transformylase in E.coli found that the reaction proceeds through a formyl phosphate intermediate. While the in vitro reaction can proceed without THF, overall the in vivo reaction is the same.[16]
Involvement in de novo Purine Biosynthesis
GART catalyzes the third step in de novo purine biosynthesis, the formation of N2-formyl-N1-(5-phospho-D-ribosyl)glycinamide (fGAR) by formyl addition to N1-(5-phospho-D-ribosyl)glycinamide (GAR).[3] In E. coli, the purN enzyme is a 23 kDa protein[17] but in humans it is part of a trifunctional protein of 110 kDa which includes AIRS and GARS functionalities.[10] This protein catalyzes three different steps of the de novo purine pathway.
Disease Relevance
Cancer Target
Due to their increased growth rate and metabolic requirements, cancer cells rely on de novo nucleotide biosynthesis to achieve levels of AMP and GMP necessary.[18] Being able to block any of the steps of the de novo purine pathway would present significant reduction in tumor growth. Studies have been done both on the substrate binding[19] and folate binding site[20] to find inhibitors.
Down Syndrome
GARTfase is suspected to be connected with Down syndrome. The gene encoding the trifunctional protein human GARS-AIRS-GART is located on chromosome 21q22.1, in the Down syndrome critical region. The protein is overexpressed in the cerebellum during the postnatal development of individuals with Down syndrome. Typically, this protein is undetectable in cerebellum shortly after birth, but found in high levels in prenatal development.[6][21]
References
- Hartman SC, Buchanan JM (July 1959). "Biosynthesis of the purines. XXVI. The identification of the formyl donors of the transformylation reactions". The Journal of Biological Chemistry. 234 (7): 1812–6. PMID 13672969.
- Smith GK, Benkovic PA, Benkovic SJ (July 1981). "L(-)-10-Formyltetrahydrofolate is the cofactor for glycinamide ribonucleotide transformylase from chicken liver". Biochemistry. 20 (14): 4034–6. doi:10.1021/bi00517a013. PMID 7284307.
- Warren L, Buchanan JM (December 1957). "Biosynthesis of the purines. XIX. 2-Amino-N-ribosylacetamide 5'-phosphate (glycinamide ribotide) transformylase". The Journal of Biological Chemistry. 229 (2): 613–26. PMID 13502326.
- McMurry, J. and Tadhg, B. The Organic Chemistry of Biological Pathways
- Connelly S, DeMartino JK, Boger DL, Wilson IA (July 2013). "Biological and structural evaluation of 10R- and 10S-methylthio-DDACTHF reveals a new role for sulfur in inhibition of glycinamide ribonucleotide transformylase". Biochemistry. 52 (30): 5133–44. doi:10.1021/bi4005182. PMC 3823235. PMID 23869564.
- Banerjee D, Nandagopal K (December 2007). "Potential interaction between the GARS-AIRS-GART Gene and CP2/LBP-1c/LSF transcription factor in Down syndrome-related Alzheimer disease". Cellular and Molecular Neurobiology. 27 (8): 1117–26. doi:10.1007/s10571-007-9217-2. PMID 17902044.
- Nygaard P, Smith JM (June 1993). "Evidence for a novel glycinamide ribonucleotide transformylase in Escherichia coli". Journal of Bacteriology. 175 (11): 3591–7. doi:10.1128/jb.175.11.3591-3597.1993. PMC 204760. PMID 8501063.
- Chen P, Schulze-Gahmen U, Stura EA, Inglese J, Johnson DL, Marolewski A, Benkovic SJ, Wilson IA (September 1992). "Crystal structure of glycinamide ribonucleotide transformylase from Escherichia coli at 3.0 A resolution. A target enzyme for chemotherapy". Journal of Molecular Biology. 227 (1): 283–92. doi:10.1016/0022-2836(92)90698-j. PMID 1522592.
- Zhang, Y., Desharnais, J., Boger, D.L., Wilson, I.A. (2005) "Human GAR Tfase complex structure with 10-(trifluoroacetyl)-5,10-dideazaacyclic-5,6,7,8-tetrahydrofolic acid and substrate beta-GAR." Unplubished. PDB: 1RBY.
- Welin M, Grossmann JG, Flodin S, Nyman T, Stenmark P, Trésaugues L, Kotenyova T, Johansson I, Nordlund P, Lehtiö L (November 2010). "Structural studies of tri-functional human GART". Nucleic Acids Research. 38 (20): 7308–19. doi:10.1093/nar/gkq595. PMC 2978367. PMID 20631005.
- Zhang Y, Desharnais J, Greasley SE, Beardsley GP, Boger DL, Wilson IA (December 2002). "Crystal structures of human GAR Tfase at low and high pH and with substrate beta-GAR". Biochemistry. 41 (48): 14206–15. doi:10.1021/bi020522m. PMID 12450384.
- Klein C, Chen P, Arevalo JH, Stura EA, Marolewski A, Warren MS, Benkovic SJ, Wilson IA (May 1995). "Towards structure-based drug design: crystal structure of a multisubstrate adduct complex of glycinamide ribonucleotide transformylase at 1.96 A resolution". Journal of Molecular Biology. 249 (1): 153–75. doi:10.1006/jmbi.1995.0286. PMID 7776369.
- Qiao QA, Jin Y, Yang C, Zhang Z, Wang M (December 2005). "A quantum chemical study on the mechanism of glycinamide ribonucleotide transformylase inhibitor: 10-Formyl-5,8,10-trideazafolic acid". Biophysical Chemistry. 118 (2–3): 78–83. doi:10.1016/j.bpc.2005.07.001. PMID 16198047.
- Inglese J, Johnson DL, Shiau A, Smith JM, Benkovic SJ (February 1990). "Subcloning, characterization, and affinity labeling of Escherichia coli glycinamide ribonucleotide transformylase". Biochemistry. 29 (6): 1436–43. doi:10.1021/bi00458a014. PMID 2185839.
- Inglese J, Smith JM, Benkovic SJ (July 1990). "Active-site mapping and site-specific mutagenesis of glycinamide ribonucleotide transformylase from Escherichia coli". Biochemistry. 29 (28): 6678–87. doi:10.1021/bi00480a018. PMID 2204419.
- Marolewski AE, Mattia KM, Warren MS, Benkovic SJ (June 1997). "Formyl phosphate: a proposed intermediate in the reaction catalyzed by Escherichia coli PurT GAR transformylase". Biochemistry. 36 (22): 6709–16. doi:10.1021/bi962961p. PMID 9184151.
- Nixon AE, Benkovic SJ (May 2000). "Improvement in the efficiency of formyl transfer of a GAR transformylase hybrid enzyme". Protein Engineering. 13 (5): 323–7. doi:10.1093/protein/13.5.323. PMID 10835105.
- Tong X, Zhao F, Thompson CB (February 2009). "The molecular determinants of de novo nucleotide biosynthesis in cancer cells". Current Opinion in Genetics & Development. 19 (1): 32–7. doi:10.1016/j.gde.2009.01.002. PMC 2707261. PMID 19201187.
- Antle VD, Donat N, Hua M, Liao PL, Vince R, Carperelli CA (October 1999). "Substrate specificity of human glycinamide ribonucleotide transformylase". Archives of Biochemistry and Biophysics. 370 (2): 231–5. doi:10.1006/abbi.1999.1428. PMID 10577357.
- Costi MP, Ferrari S (June 2001). "Update on antifolate drugs targets". Current Drug Targets. 2 (2): 135–66. doi:10.2174/1389450013348669. PMID 11469716.
- Brodsky G, Barnes T, Bleskan J, Becker L, Cox M, Patterson D (November 1997). "The human GARS-AIRS-GART gene encodes two proteins which are differentially expressed during human brain development and temporally overexpressed in cerebellum of individuals with Down syndrome". Human Molecular Genetics. 6 (12): 2043–50. doi:10.1093/hmg/6.12.2043. PMID 9328467.
External links
- Phosphoribosylglycinamide+formyltransferase at the U.S. National Library of Medicine Medical Subject Headings (MeSH)