Immunoglobulin class switching
Immunoglobulin class switching, also known as isotype switching, isotypic commutation or class-switch recombination (CSR), is a biological mechanism that changes a B cell's production of immunoglobulin from one type to another, such as from the isotype IgM to the isotype IgG.[1] During this process, the constant-region portion of the antibody heavy chain is changed, but the variable region of the heavy chain stays the same (the terms variable and constant refer to changes or lack thereof between antibodies that target different epitopes). Since the variable region does not change, class switching does not affect antigen specificity. Instead, the antibody retains affinity for the same antigens, but can interact with different effector molecules.
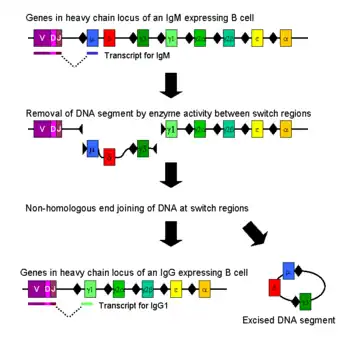
Mechanism
Class switching occurs after activation of a mature B cell via its membrane-bound antibody molecule (or B cell receptor) to generate the different classes of antibody, all with the same variable domains as the original antibody generated in the immature B cell during the process of V(D)J recombination, but possessing distinct constant domains in their heavy chains.[2]
Naïve mature B cells produce both IgM and IgD, which are the first two heavy chain segments in the immunoglobulin locus. After activation by antigen, these B cells proliferate. If these activated B cells encounter specific signaling molecules via their CD40 and cytokine receptors (both modulated by T helper cells), they undergo antibody class switching to produce IgG, IgA or IgE antibodies. During class switching, the constant region of the immunoglobulin heavy chain changes but the variable regions do not, and therefore antigenic specificity, remains the same. This allows different daughter cells from the same activated B cell to produce antibodies of different isotypes or subtypes (e.g. IgG1, IgG2 etc.).[3]
In humans, the order of the heavy chain exons is as follows:
Class switching occurs by a mechanism called class switch recombination (CSR) binding. Class switch recombination is a biological mechanism that allows the class of antibody produced by an activated B cell to change during a process known as isotype or class switching. During CSR, portions of the antibody heavy chain locus are removed from the chromosome, and the gene segments surrounding the deleted portion are rejoined to retain a functional antibody gene that produces antibody of a different isotype. Double-stranded breaks are generated in DNA at conserved nucleotide motifs, called switch (S) regions, which are upstream from gene segments that encode the constant regions of antibody heavy chains; these occur adjacent to all heavy chain constant region genes with the exception of the δ-chain. DNA is nicked and broken at two selected S-regions by the activity of a series of enzymes, including activation-induced (cytidine) deaminase (AID), uracil DNA glycosylase and apyrimidic/apurinic (AP)-endonucleases.[5][6] The intervening DNA between the S-regions is subsequently deleted from the chromosome, removing unwanted μ or δ heavy chain constant region exons and allowing substitution of a γ, α or ε constant region gene segment. The free ends of the DNA are rejoined by a process called non-homologous end joining (NHEJ) to link the variable domain exon to the desired downstream constant domain exon of the antibody heavy chain.[7] In the absence of non-homologous end joining, free ends of DNA may be rejoined by an alternative pathway biased toward microhomology joins.[8] With the exception of the μ and δ genes, only one antibody class is expressed by a B cell at any point in time. While class switch recombination is mostly a deletional process, rearranging a chromosome in "cis", it can also occur (in 10 to 20% of cases, depending upon the Ig class) as an inter-chromosomal translocation mixing immunoglobulin heavy chain genes from both alleles.[9][10]
Cytokines responsible for class switching
T cell cytokines modulate class switching in mouse (Table 1) and human (Table 2).[11][12] These cytokines may have suppressive effect on production of IgM.
Gene regulatory sequences responsible for class switching
In addition to the highly repetitive structure of the target S regions, the process of class switching needs S regions to be first transcribed and spliced out of the immunoglobulin heavy chain transcripts (where they lie within introns). Chromatin remodeling, accessibility to transcription and to AID and synapsis of broken S regions are under the control of a large super-enhancer, located downstream the more distal Calpha gene, the 3' regulatory region (3'RR).[13] In some occasions, the 3'RR super-enhancer can itself be targeted by AID and undergo DNA breaks and junction with Sμ, which then deletes the Ig heavy chain locus and defines locus suicide recombination (LSR).[14]
| T cells | Cytokines | Immunoglobulin classes | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2a | IgG2b | IgG3 | IgG4 | IgE | ||
| Th2 | IL-4 | ↑ | ↓ | ↓ | ↓ | ↓ | ↑ |
| IL-5 | ↑ | ||||||
| Th1 | IFNγ | ↓ | ↑ | ↓ | ↑ | ↓ | ↓ |
| Treg | TGFβ | ↑ | ↓ | ↑ | |||
| IL-10[15] | ↑ | ||||||
| T cells | Cytokines | Immunoglobulin classes | |||||
|---|---|---|---|---|---|---|---|
| IgG1 | IgG2 | IgG3 | IgG4 | IgA | IgE | ||
| Th2 | IL-4 | ↑ | ↓ | ↑ | ↑ | ||
| IL-5 | ↑ | ||||||
| Th1 | IFNγ | ↓ | ↑ | ↓ | |||
| Treg | TGFβ | ↓ | ↑ | ||||
| IL-10[16][17] | ↑ | ↑ | |||||
References
- Bauer G (2020). "The variability of the serological response to SARS-corona virus-2: Potential resolution of ambiguity through determination of avidity (functional affinity)". Journal of Medical Virology. 93 (1): 311–322. doi:10.1002/jmv.26262. PMC 7361859. PMID 32633840.
- Eleonora Market, F. Nina Papavasiliou (2003) V(D)J Recombination and the Evolution of the Adaptive Immune System PLoS Biology1(1): e16.
- Stavnezer J, Amemiya CT (2004). "Evolution of isotype switching". Semin. Immunol. 16 (4): 257–75. doi:10.1016/j.smim.2004.08.005. PMID 15522624.
- Parham, Peter (2015). The Immune System (4th ed.). Garland Science.
- Durandy A (2003). "Activation-induced cytidine deaminase: a dual role in class-switch recombination and somatic hypermutation". Eur. J. Immunol. 33 (8): 2069–73. doi:10.1002/eji.200324133. PMID 12884279. S2CID 32059768.
- Casali P, Zan H (2004). "Class switching and Myc translocation: how does DNA break?". Nat. Immunol. 5 (11): 1101–3. doi:10.1038/ni1104-1101. PMC 4625794. PMID 15496946.
- Lieber MR, Yu K, Raghavan SC (2006). "Roles of nonhomologous DNA end-joining, V(D)J recombination, and class switch recombination in chromosomal translocations". DNA Repair (Amst.). 5 (9–10): 1234–45. doi:10.1016/j.dnarep.2006.05.013. PMID 16793349.
- Yan CT, Boboila C, Souza EK, Franco S, Hickernell TR, Murphy M, Gumaste S, Geyer M, Zarrin AA, Manis JP, Rajewsky K, Alt FW (2007). "IgH class switching and translocations use a robust non-classical end-joining pathway". Nature. 449 (7161): 478–82. Bibcode:2007Natur.449..478Y. doi:10.1038/nature06020. PMID 17713479. S2CID 4341381.
- Reynaud S, Delpy L, Fleury L, Dougier HL, Sirac C, Cogné M (May 2005). "Interallelic class switch recombination contributes significantly to class switching in mouse B cells". Journal of Immunology. 174 (10): 6176–83. doi:10.4049/jimmunol.174.10.6176. PMID 15879114.
- Laffleur B, Bardet SM, Garot A, Brousse M, Baylet A, Cogné M (2014). "Immunoglobulin genes undergo legitimate repair in human B cells not only after cis- but also frequent trans-class switch recombination". Genes and Immunity. 15 (5): 341–6. doi:10.1038/gene.2014.25. PMID 24848929.
- Janeway CA Jr.; Travers P; Walport M; Shlomchik MJ (2001). Immunobiology (5th ed.). Garland Publishing. ISBN 978-0-8153-3642-6. (via NCBI Bookshelf).
- Male D, Brostoff J, Roth DB, Roitt I (2006). Immunology, 7th ed. Philadelphia: Mosby Elsevier, ISBN 978-0-323-03399-2 (pbk.)
- Pinaud E, Marquet M, Fiancette R, Péron S, Vincent-Fabert C, Denizot Y, Cogné M (2011). The IgH locus 3' regulatory region: pulling the strings from behind. Advances in Immunology. Vol. 110. pp. 27–70. doi:10.1016/B978-0-12-387663-8.00002-8. ISBN 9780123876638. PMID 21762815.
- Péron S, Laffleur B, Denis-Lagache N, Cook-Moreau J, Tinguely A, Delpy L, Denizot Y, Pinaud E, Cogné M (May 2012). "AID-driven deletion causes immunoglobulin heavy chain locus suicide recombination in B cells". Science. 336 (6083): 931–4. Bibcode:2012Sci...336..931P. doi:10.1126/science.1218692. PMID 22539552. S2CID 1513560.
- Shparago, N.; Zelazowski, P.; Jin, L.; McIntyre, T. M.; Stuber, E.; Peçanha, L. M.; Kehry, M. R.; Mond, J. J.; Max, E. E. (1996-05-01). "IL-10 selectively regulates murine Ig isotype switching". International Immunology. 8 (5): 781–790. doi:10.1093/intimm/8.5.781. ISSN 0953-8178. PMID 8671667.
- Brière F, Servet-Delprat C, Bridon JM, Saint-Remy JM, Banchereau J (February 1994). "Human interleukin 10 induces naive surface immunoglobulin D+ (sIgD+) B cells to secrete IgG1 and IgG3". The Journal of Experimental Medicine. 179 (2): 757–62. doi:10.1084/jem.179.2.757. PMC 2191366. PMID 8294883.
- Malisan F, Brière F, Bridon JM, Harindranath N, Mills FC, Max EE, Banchereau J, Martinez-Valdez H (March 1996). "Interleukin-10 induces immunoglobulin G isotype switch recombination in human CD40-activated naive B lymphocytes". The Journal of Experimental Medicine. 183 (3): 937–47. doi:10.1084/jem.183.3.937. PMC 2192363. PMID 8642297.
External links
- Immunoglobulin+class+switching at the U.S. National Library of Medicine Medical Subject Headings (MeSH)