Palaeolama
Palaeolama (lit. 'ancient llama') is an extinct genus of laminoid camelids that existed from the Late Pliocene to the Early Holocene (1.8 to 0.011 Ma).[2][3][4] Their range extended from North America to the intertropical region of South America.
| Palaeolama | |
|---|---|
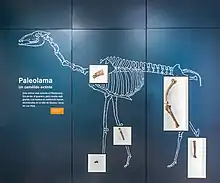 | |
| Palaeolama fossils | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Artiodactyla |
| Family: | Camelidae |
| Subfamily: | Camelinae |
| Tribe: | Lamini |
| Genus: | †Palaeolama Gervais, 1869 |
| Type species | |
| †Palaeolama weddeli Gervais, 1855 | |
| Species | |
|
†P. aequatorialis (Hoffstetter, 1952) | |
Description
Palaeolama species were relatives of modern camelids that lived in the New World from the Late Pliocene to the Late Pleistocene or Early Holocene.[2][3][4] Fossil evidence suggests that it had a slender head, elongate snout, and stocky legs.[5][6] They likely weighed around 200 kilograms (440 lb)[7] or up to 300 kilograms (660 lb), surpassing the weight of modern llamas.[6] They were specialized forest browsers and are often found in association with early equids, tapirs, deer, and mammoth.[2][4][8][9]
Morphology
Cranial
Palaeolama had a long, slender skull with an elongated rostrum and robust jaw. This morphology more closely resembles the cranial morphology of Hemiauchenia than that of modern llamas.[5]
Dental
The jaw and dental morphology of Palaeolama species distinguish them from other laminae. They tend to have a comparatively more dorsoventrally gracile mandible.[5][10] Like Hemiauchenia, Palaeolama species lack second deciduous premolars and can further be differentiated by the distinct size and shape of their third deciduous premolars. Their dentition has also been described as more brachyodont-like (short crowns, well-developed roots).[10]
Postcranial
Analyses of their limb elements reveal that they had shorter, stockier metapodials, and longer epipodials, giving them a short, stocky appearance.[5] Limbs such as these are typically associated with organisms adapted to walking on uneven and rugged terrains. This is also suggestive of being well-adapted to avoiding predators in forested areas.[4][5]
Diet
Various dietary analyses have concluded that Palaeolama was a specialized forest browser that relied almost exclusively on plants high in C3 for subsistence.[11][8][5][12] Additionally, its shallow jaw and brachydont "cheek teeth" are highly suggestive of a mixed or intermediate seasonal diet consisting of primarily leaves and fruits, with some grass.[5][11][12] Microwear analyses further validate this dietary interpretation.[5]
Group composition
As inferred from observations of modern llama, Palaeolama probably organized into bands (consisting of a single male and multiple females) and troops (consisting exclusively of young males sometimes described as "bachelors"). Typically, band territories are defended by resident males, while troops remain more or less free-roaming until they form bands of their own.[5]
Habitat

Fossil evidence suggests Palaeolama was primarily adapted to low-temperate, arid climates and preferred open, forested, and high-altitude mountainous regions.[4][9][5] The distribution of fossil evidence suggests that they had an altitudinal range limited exclusively by their dietary (vegetation) requirements.[5] Population density is shown to be highly dependent upon access and availability of subsistence resources.[5][13]
Range
The origins of this genus are a topic of much debate, as some of the earliest fossils occur during both the Irvingtonian in Florida and the Ensenadan in Uruguay.[10] Despite this, agreement exists amongst paleobiologists on the dispersal of Palaeolama during the Great American Biotic Interchange.[2][4][10]Also, some evidence suggests a move to northern South America during the second of two Pleistocene Camilidae migration events.[4] Fossil evidence ranges from the southern extent of North America (including California, Florida, and Mexico) south through Central America, and terminates in South America (Argentina and Uruguay).[2][4][9]
Palaeolama mirifica, the "stout-legged llama", is known from southern California and the Southeastern U.S., with the highest concentration of fossil specimens found in Florida (specifically the counties of Alachua, Citrus, Hillsborough, Manatee, Polk, Brevard, Orange, Sumter, and Levy). Other fossil occurrences have been discovered in Mexico, Central America (El Salvador) and South America (Argentina, Bolivia, Brazil, Chile, Colombia, Ecuador, Paraguay, Peru, Venezuela and Uruguay).[14][15]
Palaeolama major, identified by Liais in 1872, lived during the Late Pleistocene and was identified in fossil assemblages from northeastern and northern Brazil, the Pampean region of Argentina and Uruguay, northern Venezuela, and the coastal regions of Ecuador and northern Peru.[4]
Palaeolama wedelli, identified by Gervais in 1855, lived during the Mid- to Late Pleistocene, with fossil specimens found in southern Bolivia and the Andean region of Ecuador.[4]
Extinction
Climate change, changes and reductions in the types of vegetation on which they relied, and human predation are all hypothesized to have contributed to the extinction of Palaeolama during the Late Pleistocene or Early Holocene.[5][10] Evidence from both the paleoecological and fossil records suggest that Palaeolama, among other extinct camelids, weathered a number of glacial and interglacial episodes throughout their existence in North and South America. Their disappearance in some regions has been shown to coincide with a change in climate (to warmer, humid conditions) occurring at the end of the Pleistocene (also known as the Late Quaternary warming) suggesting an inability to persevere.[5][16] This hypothesis is further supported by paleoecological evidence suggesting post-megafaunal extinction shifts in vegetation and whole ecosystems.[16]
See also
References
- "Fossilworks: Palaeolama". Fossilworks. Retrieved 2020-06-05.
- Wheeler, J.C.; Chikhi, L.; Bruford, M.W. (2006). "Genetic Analysis of the Origins of Domestic South American Camelids". In Zeder, M.A.; Bradley, D.G.; Emshwiller, E.; Smith, B.D. (eds.). Documenting Domestication: New Genetic and Archaeological Paradigms. University of California Press. pp. 329–341. CiteSeerX 10.1.1.603.88. ISBN 9780520246386. JSTOR 10.1525/j.ctt1pnvs1.28.
- "Fossilworks: Palaeolama". fossilworks.org. Retrieved 2020-03-19.
- Scherer, C.S. (2013). "The Camelidae (Mammalia, Artiodactyla) from the Quaternary of South America: Cladistic and Biogeographic Hypotheses". Journal of Mammalian Evolution. 20 (1): 45–56. doi:10.1007/s10914-012-9203-4. S2CID 207195863.
- Dompierre, H. (1995). Observations on the diets of six late Cenozoic North American camelids: Camelops, Hemiauchenia, Palaeolama, Procamelus, Alforjas, and Megatylopus. National Library of Canada. ISBN 0-612-02748-1. OCLC 46500746.
- Fariña, R.A.; Vizcaíno, S.F.; De Iuliis, G. (2013). Megafauna: Giant Beasts of Pleistocene South America. Indiana University Press. ISBN 978-0-253-00230-3. JSTOR j.ctt16gzd2q. OCLC 779244424.
- Anthony J. Stuart, 2021, Vanished Giants: The Lost World of the Ice Age, "7.19 Llamas: Palaeolama and Hemiauchenia", p.136, University of Chicago Press
- Guérin, C.; Faure, M. (1999). "Palaeolama (Hemiauchenia) niedae sp. nov. from Northeastern Brazil and its place among the South American Lamini". Geobios (in French). 32 (4): 629–659. doi:10.1016/S0016-6995(99)80012-6.
- Salas, R.; Stucchi, M.; Devries, T.J. (2003). "The presence of Plio-Pleistocene Palaeolama sp. (Artiodactyla: Camelidae) on the southern coast of Peru". Bulletin de l'Institut français d'études andines. 32 (2): 347–359. doi:10.4000/bifea.6414.
- Ruez, D.R. (2005-09-30). "Earliest record of Palaeolama (Mammalia, Camelidae) with comments on "Palaeolama" guanajuatensis". Journal of Vertebrate Paleontology. 25 (3): 741–744. doi:10.1671/0272-4634(2005)025[0741:eropmc]2.0.co;2. S2CID 86522528.
- Marcolino, C.P.; Isaias, R.M. dos S.; Cozzuol, M.A.; Cartelle, C.; Dantas, M.A.T. (2012). "Diet of Palaeolama major (Camelidae) of Bahia, Brazil, inferred from coprolites". Quaternary International. 278: 81–86. Bibcode:2012QuInt.278...81M. doi:10.1016/j.quaint.2012.04.002.
- MacFadden, B.J. (2000). "Cenozoic Mammalian Herbivores From the Americas: Reconstructing Ancient Diets and Terrestrial Communities". Annual Review of Ecology and Systematics. 31 (1): 33–59. doi:10.1146/annurev.ecolsys.31.1.33.
- Ruez Jr., Dennis (September 30, 2005). "Earliest Record of Palaeolama (Mammalia, Camelidae) with Comments on "Palaeolama" guanajuatensis". Journal of Vertebrate Paleontology. 25 (3): 741–744. doi:10.1671/0272-4634(2005)025[0741:EROPMC]2.0.CO;2. JSTOR 4524496. S2CID 86522528. Retrieved 22 February 2021.
- "Fossilworks: Palaeolama mirifica". fossilworks.org.
- "'Sistine Chapel of the ancients' rock art discovered in remote Amazon forest". TheGuardian.com. 29 November 2020.
- Barnosky, .D.; Lindsey, E.L.; Villavicencio, N.A.; Bostelmann, E.; Hadly, E.A.; Wanket, J.; Marshall, C.R. (2015-10-26). "Variable impact of late-Quaternary megafaunal extinction in causing ecological state shifts in North and South America". Proceedings of the National Academy of Sciences. 113 (4): 856–861. doi:10.1073/pnas.1505295112. PMC 4739530. PMID 26504219.