Multisensory integration
Multisensory integration, also known as multimodal integration, is the study of how information from the different sensory modalities (such as sight, sound, touch, smell, self-motion, and taste) may be integrated by the nervous system.[1] A coherent representation of objects combining modalities enables animals to have meaningful perceptual experiences. Indeed, multisensory integration is central to adaptive behavior because it allows animals to perceive a world of coherent perceptual entities.[2] Multisensory integration also deals with how different sensory modalities interact with one another and alter each other's processing.
General introduction
Multimodal perception is how animals form coherent, valid, and robust perception by processing sensory stimuli from various modalities. Surrounded by multiple objects and receiving multiple sensory stimulations, the brain is faced with the decision of how to categorize the stimuli resulting from different objects or events in the physical world. The nervous system is thus responsible for whether to integrate or segregate certain groups of signals. Multimodal perception has been widely studied in cognitive science, behavioral science, and neuroscience.
Stimuli and sensory modalities
There are four attributes of stimulus: modality, intensity, location, and duration. The neocortex in the mammalian brain has parcellations that primarily process sensory input from one modality. For example, primary visual area, V1, or primary somatosensory area, S1. These areas mostly deal with low-level stimulus features such as brightness, orientation, intensity, etc. These areas have extensive connections to each other as well as to higher association areas that further process the stimuli and are believed to integrate sensory input from various modalities. However, multisensory effects have been shown to occur in primary sensory areas as well.[3]
Binding problem
The relationship between the binding problem and multisensory perception can be thought of as a question – the binding problem – and its potential solution – multisensory perception. The binding problem stemmed from unanswered questions about how mammals (particularly higher primates) generate a unified, coherent perception of their surroundings from the cacophony of electromagnetic waves, chemical interactions, and pressure fluctuations that forms the physical basis of the world around us. It was investigated initially in the visual domain (colour, motion, depth, and form), then in the auditory domain, and recently in the multisensory areas. It can be said therefore, that the binding problem is central to multisensory perception.[4]
However, considerations of how unified conscious representations are formed are not the full focus of multisensory Integration research. It is obviously important for the senses to interact in order to maximize how efficiently people interact with the environment. For perceptual experience and behavior to benefit from the simultaneous stimulation of multiple sensory modalities, integration of the information from these modalities is necessary. Some of the mechanisms mediating this phenomenon and its subsequent effects on cognitive and behavioural processes will be examined hereafter. Perception is often defined as one's conscious experience, and thereby combines inputs from all relevant senses and prior knowledge. Perception is also defined and studied in terms of feature extraction, which is several hundred milliseconds away from conscious experience. Notwithstanding the existence of Gestalt psychology schools that advocate a holistic approach to the operation of the brain,[5][6] the physiological processes underlying the formation of percepts and conscious experience have been vastly understudied. Nevertheless, burgeoning neuroscience research continues to enrich our understanding of the many details of the brain, including neural structures implicated in multisensory integration such as the superior colliculus (SC)[7] and various cortical structures such as the superior temporal gyrus (GT) and visual and auditory association areas. Although the structure and function of the SC are well known, the cortex and the relationship between its constituent parts are presently the subject of much investigation. Concurrently, the recent impetus on integration has enabled investigation into perceptual phenomena such as the ventriloquism effect,[8] rapid localization of stimuli and the McGurk effect;[9] culminating in a more thorough understanding of the human brain and its functions.
History
Studies of sensory processing in humans and other animals has traditionally been performed one sense at a time,[10] and to the present day, numerous academic societies and journals are largely restricted to considering sensory modalities separately ('Vision Research', 'Hearing Research' etc.). However, there is also a long and parallel history of multisensory research. An example is the Stratton's (1896) experiments on the somatosensory effects of wearing vision-distorting prism glasses.[11][12] Multisensory interactions or crossmodal effects in which the perception of a stimulus is influenced by the presence of another type of stimulus are referred since very early in the past. They were reviewed by Hartmann[13] in a fundamental book where, among several references to different types of multisensory interactions, reference is made to the work of Urbantschitsch in 1888[14] who reported on the improvement of visual acuity by auditive stimuli in subjects with damaged brains. This effect was also found later in individuals with undamaged brains by Krakov[15] and Hartmann,[16] as well as the fact that the visual acuity could be improved by other type of stimuli.[16] It is also noteworthy the amount of work in the early 1930s on intersensory relations in the Soviet Union, reviewed by London.[17] A remarkable multisensory research is the extensive work of Gonzalo [18] in the forties on the characterization of a multisensory syndrome in patients with parieto-occipital cortical lesions. In this syndrome, all the sensory functions are affected, and with symmetric bilaterality, in spite of being a unilateral lesion where the primary areas were not involved. A feature of this syndrome is the great permeability to crossmodal effects between visual, tactile, auditive stimuli as well as muscular effort to improve the perception, also decreasing the reaction times. The improvement by crossmodal effect was found to be greater as the primary stimulus to be perceived was weaker, and as the cortical lesion was greater (Vol I and II of reference[18]). This author interpreted these phenomena under a dynamic physiological concept, and from a model based on functional gradients through the cortex and scaling laws of dynamical systems, thus highlighting the functional unity of the cortex. According to the functional cortical gradients, the specificity of the cortex would be distributed in gradation, and the overlap of different specific gradients would be related to multisensory interactions.[19]
Multisensory research has recently gained enormous interest and popularity.
Example of spatial and structural congruence
When we hear a car honk, we would determine which car triggers the honk by which car we see is the spatially closest to the honk. It's a spatially congruent example by combining visual and auditory stimuli. On the other hand, the sound and the pictures of a TV program would be integrated as structurally congruent by combining visual and auditory stimuli. However, if the sound and the pictures did not meaningfully fit, we would segregate the two stimuli. Therefore, spatial or structural congruence comes from not only combining the stimuli but is also determined by our understanding.
Theories and approaches
Visual dominance
Literature on spatial crossmodal biases suggests that visual modality often influences information from other senses.[20] Some research indicates that vision dominates what we hear, when varying the degree of spatial congruency. This is known as the ventriloquist effect.[21] In cases of visual and haptic integration, children younger than 8 years of age show visual dominance when required to identify object orientation. However, haptic dominance occurs when the factor to identify is object size.[22][23]
Modality appropriateness
According to Welch and Warren (1980), the Modality Appropriateness Hypothesis states that the influence of perception in each modality in multisensory integration depends on that modality's appropriateness for the given task. Thus, vision has a greater influence on integrated localization than hearing, and hearing and touch have a greater bearing on timing estimates than vision.[24][25]
More recent studies refine this early qualitative account of multisensory integration. Alais and Burr (2004), found that following progressive degradation in the quality of a visual stimulus, participants' perception of spatial location was determined progressively more by a simultaneous auditory cue.[26] However, they also progressively changed the temporal uncertainty of the auditory cue; eventually concluding that it is the uncertainty of individual modalities that determine to what extent information from each modality is considered when forming a percept.[26] This conclusion is similar in some respects to the 'inverse effectiveness rule'. The extent to which multisensory integration occurs may vary according to the ambiguity of the relevant stimuli. In support of this notion, a recent study shows that weak senses such as olfaction can even modulate the perception of visual information as long as the reliability of visual signals is adequately compromised.[27]
Bayesian integration
The theory of Bayesian integration is based on the fact that the brain must deal with a number of inputs, which vary in reliability.[28] In dealing with these inputs, it must construct a coherent representation of the world that corresponds to reality. The Bayesian integration view is that the brain uses a form of Bayesian inference.[29] This view has been backed up by computational modeling of such a Bayesian inference from signals to coherent representation, which shows similar characteristics to integration in the brain.[29]
Cue combination vs. causal inference models
With the assumption of independence between various sources, the traditional cue combination model is successful in modality integration. However, depending on the discrepancies between modalities, there might be different forms of stimuli fusion: integration, partial integration, and segregation. To fully understand the other two types, we have to use causal inference model without the assumption as cue combination model. This freedom gives us general combination of any numbers of signals and modalities by using Bayes' rule to make causal inference of sensory signals.[30]
The hierarchical vs. non-hierarchical models
The difference between two models is that hierarchical model can explicitly make causal inference to predict certain stimulus while non-hierarchical model can only predict joint probability of stimuli. However, hierarchical model is actually a special case of non-hierarchical model by setting joint prior as a weighted average of the prior to common and independent causes, each weighted by their prior probability. Based on the correspondence of these two models, we can also say that hierarchical is a mixture modal of non-hierarchical model.
Independence of likelihoods and priors
For Bayesian model, the prior and likelihood generally represent the statistics of the environment and the sensory representations. The independence of priors and likelihoods is not assured since the prior may vary with likelihood only by the representations. However, the independence has been proved by Shams with series of parameter control in multi sensory perception experiment.[31]
Principles
The contributions of Barry Stein, Alex Meredith, and their colleagues (e.g."The merging of the senses" 1993,[32]) are widely considered to be the groundbreaking work in the modern field of multisensory integration. Through detailed long-term study of the neurophysiology of the superior colliculus, they distilled three general principles by which multisensory integration may best be described.
- The spatial rule[33][34] states that multisensory integration is more likely or stronger when the constituent unisensory stimuli arise from approximately the same location.
- The temporal rule[34][35] states that multisensory integration is more likely or stronger when the constituent unisensory stimuli arise at approximately the same time.
- The principle of inverse effectiveness[36][37] states that multisensory integration is more likely or stronger when the constituent unisensory stimuli evoke relatively weak responses when presented in isolation.
Perceptual and behavioral consequences
A unimodal approach dominated scientific literature until the beginning of this century. Although this enabled rapid progression of neural mapping, and an improved understanding of neural structures, the investigation of perception remained relatively stagnant, with a few exceptions. The recent revitalized enthusiasm into perceptual research is indicative of a substantial shift away from reductionism and toward gestalt methodologies. Gestalt theory, dominant in the late 19th and early 20th centuries espoused two general principles: the 'principle of totality' in which conscious experience must be considered globally, and the 'principle of psychophysical isomorphism' which states that perceptual phenomena are correlated with cerebral activity. Just these ideas were already applied by Justo Gonzalo in his work of brain dynamics, where a sensory-cerebral correspondence is considered in the formulation of the "development of the sensory field due to a psychophysical isomorphism" (pag. 23 of the English translation of ref.[19]). Both ideas 'principle of totality' and 'psychophysical isomorphism' are particularly relevant in the current climate and have driven researchers to investigate the behavioural benefits of multisensory integration.
Decreasing sensory uncertainty
It has been widely acknowledged that uncertainty in sensory domains results in an increased dependence of multisensory integration.[26] Hence, it follows that cues from multiple modalities that are both temporally and spatially synchronous are viewed neurally and perceptually as emanating from the same source. The degree of synchrony that is required for this 'binding' to occur is currently being investigated in a variety of approaches.[38] The integrative function only occurs to a point beyond which the subject can differentiate them as two opposing stimuli. Concurrently, a significant intermediate conclusion can be drawn from the research thus far. Multisensory stimuli that are bound into a single percept, are also bound on the same receptive fields of multisensory neurons in the SC and cortex.[26]
Decreasing reaction time
Responses to multiple simultaneous sensory stimuli can be faster than responses to the same stimuli presented in isolation. Hershenson (1962) presented a light and tone simultaneously and separately, and asked human participants to respond as rapidly as possible to them. As the asynchrony between the onsets of both stimuli was varied, it was observed that for certain degrees of asynchrony, reaction times were decreased.[39] These levels of asynchrony were quite small, perhaps reflecting the temporal window that exists in multisensory neurons of the SC. Further studies have analysed the reaction times of saccadic eye movements;[40] and more recently correlated these findings to neural phenomena.[41] In patients studied by Gonzalo,[18] with lesions in the parieto-occipital cortex, the decrease in the reaction time to a given stimulus by means of intersensory facilitation was shown to be very remarkable.
Redundant target effects
The redundant target effect is the observation that people typically respond faster to double targets (two targets presented simultaneously) than to either of the targets presented alone. This difference in latency is termed the redundancy gain (RG).[42]
In a study done by Forster, Cavina-Pratesi, Aglioti, and Berlucchi (2001), normal observers responded faster to simultaneous visual and tactile stimuli than to single visual or tactile stimuli. RT to simultaneous visual and tactile stimuli was also faster than RT to simultaneous dual visual or tactile stimuli. The advantage for RT to combined visual-tactile stimuli over RT to the other types of stimulation could be accounted for by intersensory neural facilitation rather than by probability summation. These effects can be ascribed to the convergence of tactile and visual inputs onto neural centers which contain flexible multisensory representations of body parts.[43]
McGurk effect
It has been found that two converging bimodal stimuli can produce a perception that is not only different in magnitude than the sum of its parts, but also quite different in quality. In a classic study labeled the McGurk effect,[44] a person's phoneme production was dubbed with a video of that person speaking a different phoneme.[45] The result was the perception of a third, different phoneme. McGurk and MacDonald (1976) explained that phonemes such as ba, da, ka, ta, ga and pa can be divided into four groups, those that can be visually confused, i.e. (da, ga, ka, ta) and (ba and pa), and those that can be audibly confused. Hence, when ba – voice and ga lips are processed together, the visual modality sees ga or da, and the auditory modality hears ba or da, combining to form the percept da.[44]
Ventriloquism
Ventriloquism has been used as the evidence for the modality appropriateness hypothesis. The ventriloquism effect is the situation in which auditory location perception is shifted toward a visual cue. The original study describing this phenomenon was conducted by Howard and Templeton, (1966) after which several studies have replicated and built upon the conclusions they reached.[46] In conditions in which the visual cue is unambiguous, visual capture reliably occurs. Thus to test the influence of sound on perceived location, the visual stimulus must be progressively degraded.[26] Furthermore, given that auditory stimuli are more attuned to temporal changes, recent studies have tested the ability of temporal characteristics to influence the spatial location of visual stimuli. Some types of EVP – electronic voice phenomenon, mainly the ones using sound bubbles are considered a kind of modern ventriloquism technique and is played by the use of sophisticated software, computers and sound equipment.
Double-flash illusion
The double flash illusion was reported as the first illusion to show that visual stimuli can be qualitatively altered by audio stimuli.[47] In the standard paradigm participants are presented combinations of one to four flashes accompanied by zero to 4 beeps. They were then asked to say how many flashes they perceived. Participants perceived illusory flashes when there were more beeps than flashes. fMRI studies have shown that there is crossmodal activation in early, low level visual areas, which was qualitatively similar to the perception of a real flash. This suggests that the illusion reflects subjective perception of the extra flash.[48] Further, studies suggest that timing of multisensory activation in unisensory cortexes is too fast to be mediated by a higher order integration suggesting feed forward or lateral connections.[49] One study has revealed the same effect but from vision to audition, as well as fission rather than fusion effects, although the level of the auditory stimulus was reduced to make it less salient for those illusions affecting audition.[50]
Rubber hand illusion
In the rubber hand illusion (RHI),[51] human participants view a dummy hand being stroked with a paintbrush, while they feel a series of identical brushstrokes applied to their own hand, which is hidden from view. If this visual and tactile information is applied synchronously, and if the visual appearance and position of the dummy hand is similar to one's own hand, then people may feel that the touches on their own hand are coming from the dummy hand, and even that the dummy hand is, in some way, their own hand.[51] This is an early form of body transfer illusion. The RHI is an illusion of vision, touch, and posture (proprioception), but a similar illusion can also be induced with touch and proprioception.[52] It has also been found that the illusion may not require tactile stimulation at all, but can be completely induced using mere vision of the rubber hand being in a congruent posture with the hidden real hand.[53] The very first report of this kind of illusion may have been as early as 1937 (Tastevin, 1937).[54][55][56]
Body transfer illusion
Body transfer illusion typically involves the use of virtual reality devices to induce the illusion in the subject that the body of another person or being is the subject's own body.
Neural mechanisms
Superior colliculus
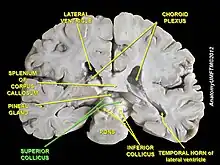
The superior colliculus (SC) or optic tectum (OT) is part of the tectum, located in the midbrain, superior to the brainstem and inferior to the thalamus. It contains seven layers of alternating white and grey matter, of which the superficial contain topographic maps of the visual field; and deeper layers contain overlapping spatial maps of the visual, auditory and somatosensory modalities.[57] The structure receives afferents directly from the retina, as well as from various regions of the cortex (primarily the occipital lobe), the spinal cord and the inferior colliculus. It sends efferents to the spinal cord, cerebellum, thalamus and occipital lobe via the lateral geniculate nucleus (LGN). The structure contains a high proportion of multisensory neurons and plays a role in the motor control of orientation behaviours of the eyes, ears and head.[41]
Receptive fields from somatosensory, visual and auditory modalities converge in the deeper layers to form a two-dimensional multisensory map of the external world. Here, objects straight ahead are represented caudally and objects on the periphery are represented rosterally. Similarly, locations in superior sensory space are represented medially, and inferior locations are represented laterally.[32]
However, in contrast to simple convergence, the SC integrates information to create an output that differs from the sum of its inputs. Following a phenomenon labelled the 'spatial rule', neurons are excited if stimuli from multiple modalities fall on the same or adjacent receptive fields, but are inhibited if the stimuli fall on disparate fields.[58] Excited neurons may then proceed to innervate various muscles and neural structures to orient an individual's behaviour and attention toward the stimulus. Neurons in the SC also adhere to the 'temporal rule', in which stimulation must occur within close temporal proximity to excite neurons. However, due to the varying processing time between modalities and the relatively slower speed of sound to light, it has been found the neurons may be optimally excited when stimulated some time apart.[59]
Putamen
Single neurons in the macaque putamen have been shown to have visual and somatosensory responses closely related to those in the polysensory zone of the premotor cortex and area 7b in the parietal lobe.[60][61]
Cortical areas
Multisensory neurons exist in a large number of locations, often integrated with unimodal neurons. They have recently been discovered in areas previously thought to be modality specific, such as the somatosensory cortex; as well as in clusters at the borders between the major cerebral lobes, such as the occipito-parietal space and the occipito-temporal space.[62][41][63]
However, in order to undergo such physiological changes, there must exist continuous connectivity between these multisensory structures. It is generally agreed that information flow within the cortex follows a hierarchical configuration.[64] Hubel and Wiesel showed that receptive fields and thus the function of cortical structures, as one proceeds out from V1 along the visual pathways, become increasingly complex and specialized.[64] From this it was postulated that information flowed outwards in a feed-forward fashion; the complex end products eventually binding to form a percept. However, via fMRI and intracranial recording technologies, it has been observed that the activation time of successive levels of the hierarchy does not correlate with a feed-forward structure. That is, late activation has been observed in the striate cortex, markedly after activation of the prefrontal cortex in response to the same stimulus.[65]
Complementing this, afferent nerve fibres have been found that project to early visual areas such as the lingual gyrus from late in the dorsal (action) and ventral (perception) visual streams, as well as from the auditory association cortex.[66] Feedback projections have also been observed in the opossum directly from the auditory association cortex to V1.[64] This last observation currently highlights a point of controversy within the neuroscientific community. Sadato et al. (2004) concluded, in line with Bernstein et al. (2002), that the primary auditory cortex (A1) was functionally distinct from the auditory association cortex, in that it was void of any interaction with the visual modality. They hence concluded that A1 would not at all be effected by cross modal plasticity.[67][68] This concurs with Jones and Powell's (1970) contention that primary sensory areas are connected only to other areas of the same modality.[69]
In contrast, the dorsal auditory pathway, projecting from the temporal lobe is largely concerned with processing spatial information, and contains receptive fields that are topographically organized. Fibers from this region project directly to neurons governing corresponding receptive fields in V1.[64] The perceptual consequences of this have not yet been empirically acknowledged. However, it can be hypothesized that these projections may be the precursors of increased acuity and emphasis of visual stimuli in relevant areas of perceptual space. Consequently, this finding rejects Jones and Powell's (1970) hypothesis[69] and thus is in conflict with Sadato et al.'s (2004) findings.[67] A resolution to this discrepancy includes the possibility that primary sensory areas can not be classified as a single group, and thus may be far more different from what was previously thought.
The multisensory syndrome with symmetric bilaterality, characterized by Gonzalo and called by this author 'central syndrome of the cortex',[18][19] was originated from a unilateral parieto-occipital cortical lesion equidistant from the visual, tactile, and auditory projection areas (the middle of area 19, the anterior part of area 18 and the most posterior of area 39, in Brodmann terminology) that was called 'central zone'. The gradation observed between syndromes led this author to propose a functional gradient scheme in which the specificity of the cortex is distributed with a continuous variation,[19] the overlap of the specific gradients would be high or maximum in that 'central zone'.
Occipital lobe
Primary visual cortex (V1)[73]
Lingual gyrus in humans
Lateral occipital complex (LOC), including lateral occipital tactile visual area (LOtv)[74]
Parietal lobe
Ventral intraparietal sulcus (VIP) in macaques[70]
Lateral intraparietal sulcus (LIP) in macaques[70]
Area 7b in macaques[75]
Second somatosensory cortex (SII)[76]
Temporal lobe
Primary auditory cortex (A1)
Superior temporal cortex (STG/STS/PT) Audio visual cross modal interactions are known to occur in the auditory association cortex which lies directly inferior to the Sylvian fissure in the temporal lobe.[67] Plasticity was observed in the superior temporal gyrus (STG) by Petitto et al. (2000).[77] Here, it was found that the STG was more active during stimulation in native deaf signers compared to hearing non signers. Concurrently, further research has revealed differences in the activation of the Planum temporale (PT) in response to non linguistic lip movements between the hearing and deaf; as well as progressively increasing activation of the auditory association cortex as previously deaf participants gain hearing experience via a cochlear implant.[67]
Anterior ectosylvian sulus (AES) in cats[78][79][80]
Rostral lateral suprasylvian sulcus (rLS) in cats[79]
Cortical-subcortical interactions
The most significant interaction between these two systems (corticotectal interactions) is the connection between the anterior ectosylvian sulcus (AES), which lies at the junction of the parietal, temporal and frontal lobes, and the SC. The AES is divided into three unimodal regions with multisensory neurons at the junctions between these sections.[81] (Jiang & Stein, 2003). Neurons from the unimodal regions project to the deep layers of the SC and influence the multiplicative integration effect. That is, although they can receive inputs from all modalities as normal, the SC can not enhance or depress the effect of multisensory stimulation without input from the AES.[81]
Concurrently, the multisensory neurons of the AES, although also integrally connected to unimodal AES neurons, are not directly connected to the SC. This pattern of division is reflected in other areas of the cortex, resulting in the observation that cortical and tectal multisensory systems are somewhat dissociated.[82] Stein, London, Wilkinson and Price (1996) analysed the perceived luminance of an LED in the context of spatially disparate auditory distracters of various types. A significant finding was that a sound increased the perceived brightness of the light, regardless of their relative spatial locations, provided the light's image was projected onto the fovea.[83] Here, the apparent lack of the spatial rule, further differentiates cortical and tectal multisensory neurons. Little empirical evidence exists to justify this dichotomy. Nevertheless, cortical neurons governing perception, and a separate sub cortical system governing action (orientation behavior) is synonymous with the perception action hypothesis of the visual stream.[84] Further investigation into this field is necessary before any substantial claims can be made.
Dual "what" and "where" multisensory routes
Research suggests the existence of two multisensory routes for "what" and "where". The "what" route identifying the identity of things involving area Brodmann area 9 in the right inferior frontal gyrus and right middle frontal gyrus, Brodmann area 13 and Brodmann area 45 in the right insula-inferior frontal gyrus area, and Brodmann area 13 bilaterally in the insula. The "where" route detecting their spatial attributes involving the Brodmann area 40 in the right and left inferior parietal lobule and the Brodmann area 7 in the right precuneus-superior parietal lobule and Brodmann area 7 in the left superior parietal lobule.[85]
Development of multisensory operations
Theories of development
All species equipped with multiple sensory systems, utilize them in an integrative manner to achieve action and perception.[32] However, in most species, especially higher mammals and humans, the ability to integrate develops in parallel with physical and cognitive maturity. Children until certain ages do not show mature integration patterns.[86][87] Classically, two opposing views that are principally modern manifestations of the nativist/empiricist dichotomy have been put forth. The integration (empiricist) view states that at birth, sensory modalities are not at all connected. Hence, it is only through active exploration that plastic changes can occur in the nervous system to initiate holistic perceptions and actions. Conversely, the differentiation (nativist) perspective asserts that the young nervous system is highly interconnected; and that during development, modalities are gradually differentiated as relevant connections are rehearsed and the irrelevant are discarded.[88]
Using the SC as a model, the nature of this dichotomy can be analysed. In the newborn cat, deep layers of the SC contain only neurons responding to the somatosensory modality. Within a week, auditory neurons begin to occur, but it is not until two weeks after birth that the first multisensory neurons appear. Further changes continue, with the arrival of visual neurons after three weeks, until the SC has achieved its fully mature structure after three to four months. Concurrently in species of monkey, newborns are endowed with a significant complement of multisensory cells; however, along with cats there is no integration effect apparent until much later.[41] This delay is thought to be the result of the relatively slower development of cortical structures including the AES; which as stated above, is essential for the existence of the integration effect.[81]
Furthermore, it was found by Wallace (2004) that cats raised in a light deprived environment had severely underdeveloped visual receptive fields in deep layers of the SC.[41] Although, receptive field size has been shown to decrease with maturity, the above finding suggests that integration in the SC is a function of experience. Nevertheless, the existence of visual multisensory neurons, despite a complete lack of visual experience, highlights the apparent relevance of nativist viewpoints. Multisensory development in the cortex has been studied to a lesser extent, however a similar study to that presented above was performed on cats whose optic nerves had been severed. These cats displayed a marked improvement in their ability to localize stimuli through audition; and consequently also showed increased neural connectivity between V1 and the auditory cortex.[64] Such plasticity in early childhood allows for greater adaptability, and thus more normal development in other areas for those with a sensory deficit.
In contrast, following the initial formative period, the SC does not appear to display any neural plasticity. Despite this, habituation and sensititisation over the long term is known to exist in orientation behaviors. This apparent plasticity in function has been attributed to the adaptability of the AES. That is, although neurons in the SC have a fixed magnitude of output per unit input, and essentially operate an all or nothing response, the level of neural firing can be more finely tuned by variations in input by the AES.
Although there is evidence for either perspective of the integration/differentiation dichotomy, a significant body of evidence also exists for a combination of factors from either view. Thus, analogous to the broader nativist/empiricist argument, it is apparent that rather than a dichotomy, there exists a continuum, such that the integration and differentiation hypotheses are extremes at either end.
Psychophysical development of integration
Not much is known about the development of the ability to integrate multiple estimates such as vision and touch.[86] Some multisensory abilities are present from early infancy, but it is not until children are eight years or older before they use multiple modalities to reduce sensory uncertainty.[86]
One study demonstrated that cross-modal visual and auditory integration is present from within 1 year of life.[89] This study measured response time for orientating towards a source. Infants who were 8–10 months old showed significantly decreased response times when the source was presented through both visual and auditory information compared to a single modality. Younger infants, however, showed no such change in response times to these different conditions. Indeed, the results of the study indicates that children potentially have the capacity to integrate sensory sources at any age. However, in certain cases, for example visual cues, intermodal integration is avoided.[86]
Another study found that cross-modal integration of touch and vision for distinguishing size and orientation is available from at least 8 years of age.[87] For pre-integration age groups, one sense dominates depending on the characteristic discerned (see visual dominance).[87]
A study investigating sensory integration within a single modality (vision) found that it cannot be established until age 12 and above.[86] This particular study assessed the integration of disparity and texture cues to resolve surface slant. Though younger age groups showed a somewhat better performance when combining disparity and texture cues compared to using only disparity or texture cues, this difference was not statistically significant.[86] In adults, the sensory integration can be mandatory, meaning that they no longer have access to the individual sensory sources.[90]
Acknowledging these variations, many hypotheses have been established to reflect why these observations are task-dependent. Given that different senses develop at different rates, it has been proposed that cross-modal integration does not appear until both modalities have reached maturity.[87][91] The human body undergoes significant physical transformation throughout childhood. Not only is there growth in size and stature (affecting viewing height), but there is also change in inter-ocular distance and eyeball length. Therefore, sensory signals need to be constantly re-evaluated to appreciate these various physiological changes.[87] Some support comes from animal studies that explore the neurobiology behind integration. Adult monkeys have deep inter-neuronal connections within the superior colliculus providing strong, accelerated visuo-auditory integration.[92] Young animals conversely, do not have this enhancement until unimodal properties are fully developed.[93][94]
Additionally, to rationalize sensory dominance, Gori et al. (2008) advocates that the brain utilises the most direct source of information during sensory immaturity.[87] In this case, orientation is primarily a visual characteristic. It can be derived directly from the object image that forms on the retina, irrespective of other visual factors. In fact, data shows that a functional property of neurons within primate visual cortices' are their discernment to orientation.[95] In contrast, haptic orientation judgements are recovered through collaborated patterned stimulations, evidently an indirect source susceptible to interference. Likewise, when size is concerned haptic information coming from positions of the fingers is more immediate. Visual-size perceptions, alternatively, have to be computed using parameters such as slant and distance. Considering this, sensory dominance is a useful instinct to assist with calibration. During sensory immaturity, the more simple and robust information source could be used to tweak the accuracy of the alternate source.[87] Follow-up work by Gori et al. (2012) showed that, at all ages, vision-size perceptions are near perfect when viewing objects within the haptic workspace (i.e. at arm's reach).[96] However, systematic errors in perception appeared when the object was positioned beyond this zone.[97] Children younger than 14 years tend to underestimate object size, whereas adults overestimated. However, if the object was returned to the haptic workspace, those visual biases disappeared.[96] These results support the hypothesis that haptic information may educate visual perceptions. If sources are used for cross-calibration they cannot, therefore, be combined (integrated). Maintaining access to individual estimates is a trade-off for extra plasticity over accuracy, which could be beneficial in retrospect to the developing body.[87][91]
Alternatively, Ernst (2008) advocates that efficient integration initially relies upon establishing correspondence – which sensory signals belong together.[91] Indeed, studies have shown that visuo-haptic integration fails in adults when there is a perceived spatial separation, suggesting sensory information is coming from different targets.[98] Furthermore, if the separation can be explained, for example viewing an object through a mirror, integration is re-established and can even be optimal.[99][100] Ernst (2008) suggests that adults can obtain this knowledge from previous experiences to quickly determine which sensory sources depict the same target, but young children could be deficient in this area.[91] Once there is a sufficient bank of experiences, confidence to correctly integrate sensory signals can then be introduced in their behaviour.
Lastly, Nardini et al. (2010) recently hypothesised that young children have optimized their sensory appreciation for speed over accuracy.[86] When information is presented in two forms, children may derive an estimate from the fastest available source, subsequently ignoring the alternate, even if it contains redundant information. Nardini et al. (2010) provides evidence that children's (aged 6 years) response latencies are significantly lower when stimuli are presented in multi-cue over single-cue conditions.[86] Conversely, adults showed no change between these conditions. Indeed, adults display mandatory fusion of signals, therefore they can only ever aim for maximum accuracy.[86][90] However, the overall mean latencies for children were not faster than adults, which suggests that speed optimization merely enable them to keep up with the mature pace. Considering the haste of real-world events, this strategy may prove necessary to counteract the general slower processing of children and maintain effective vision-action coupling.[101][102][103] Ultimately the developing sensory system may preferentially adapt for different goals – speed and detecting sensory conflicts – those typical of objective learning.
The late development of efficient integration has also been investigated from computational point of view.[104] Daee et al. (2014) showed that having one dominant sensory source at early age, rather than integrating all sources, facilitates the overall development of cross-modal integrations.
Prosthesis
Prosthetics designers should carefully consider the nature of dimensionality alteration of sensorimotor signaling from and to the CNS when designing prosthetic devices. As reported in literatures, neural signaling from the CNS to the motors is organized in a way that the dimensionalities of the signals are gradually increased as you approach the muscles, also called muscle synergies. In the same principal, but in opposite ordering, on the other hand, signals dimensionalities from the sensory receptors are gradually integrated, also called sensory synergies, as they approaches the CNS. This bow tie like signaling formation enables the CNS to process abstract yet valuable information only. Such as process will decrease complexity of the data, handle the noises and guarantee to the CNS the optimum energy consumption. Although the current commercially available prosthetic devices mainly focusing in implementing the motor side by simply uses EMG sensors to switch between different activation states of the prosthesis. Very limited works have proposed a system to involve by integrating the sensory side. The integration of tactile sense and proprioception is regarded as essential for implementing the ability to perceive environmental input.[105]
Visual Rehabilitation
Multisensory integration has also been shown to ameliorate visual hemianopia. Through the repeated presentation of multisensory stimuli in the blind hemifield, the ability to respond to purely visual stimuli gradually returns to that hemifield in a central to peripheral manner. These benefits persist even after the explicit multisensory training ceases.[106]
References
- Stein, BE.; Stanford, TR.; Rowland, BA. (Dec 2009). "The neural basis of multisensory integration in the midbrain: its organization and maturation". Hear Res. 258 (1–2): 4–15. doi:10.1016/j.heares.2009.03.012. PMC 2787841. PMID 19345256.
- Lewkowicz DJ, Ghazanfar AA (November 2009). "The emergence of multisensory systems through perceptual narrowing" (PDF). Trends Cogn. Sci. (Regul. Ed.). 13 (11): 470–8. CiteSeerX 10.1.1.554.4323. doi:10.1016/j.tics.2009.08.004. PMID 19748305. S2CID 14289579.
- Lemus L, Hernández A, Luna R, Zainos A, Romo R (July 2010). "Do sensory cortices process more than one sensory modality during perceptual judgements?". Neuron. 67 (2): 335–48. doi:10.1016/j.neuron.2010.06.015. PMID 20670839. S2CID 16043442.
- Zmigrod, S.; Hommel, B. (Jan 2010). "Temporal dynamics of unimodal and multimodal feature binding" (PDF). Atten Percept Psychophys. 72 (1): 142–52. doi:10.3758/APP.72.1.142. PMID 20045885. S2CID 7055915.
- Wagemans, J.; Elder, JH.; Kubovy, M.; Palmer, SE.; Peterson, MA.; Singh, M.; von der Heydt, R. (Nov 2012). "A century of Gestalt psychology in visual perception: I. Perceptual grouping and figure-ground organization". Psychol Bull. 138 (6): 1172–217. CiteSeerX 10.1.1.452.8394. doi:10.1037/a0029333. PMC 3482144. PMID 22845751.
- Wagemans, J.; Feldman, J.; Gepshtein, S.; Kimchi, R.; Pomerantz, JR.; van der Helm, PA.; van Leeuwen, C. (Nov 2012). "A century of Gestalt psychology in visual perception: II. Conceptual and theoretical foundations". Psychol Bull. 138 (6): 1218–52. doi:10.1037/a0029334. PMC 3728284. PMID 22845750.
- Stein, BE.; Rowland, BA. (2011). "Organization and plasticity in multisensory integration". Enhancing Performance for Action and Perception - Multisensory Integration, Neuroplasticity and Neuroprosthetics, Part I. pp. 145–63. doi:10.1016/B978-0-444-53752-2.00007-2. ISBN 9780444537522. PMC 3245961. PMID 21741550.
{{cite book}}:|journal=ignored (help) - Recanzone, GH. (Dec 2009). "Interactions of auditory and visual stimuli in space and time". Hear Res. 258 (1–2): 89–99. doi:10.1016/j.heares.2009.04.009. PMC 2787663. PMID 19393306.
- Smith, E.; Duede, S.; Hanrahan, S.; Davis, T.; House, P.; Greger, B. (2013). "Seeing is believing: neural representations of visual stimuli in human auditory cortex correlate with illusory auditory perceptions". PLOS ONE. 8 (9): e73148. Bibcode:2013PLoSO...873148S. doi:10.1371/journal.pone.0073148. PMC 3762867. PMID 24023823.
- Fodor, Jerry A. (1983). Modularity of mind: an essay on faculty psychology. Cambridge, Mass: MIT Press. ISBN 978-0-262-06084-4. OCLC 551957787.
- Stratton, George M. (1896). "Some preliminary experiments on vision without inversion of the retinal image". Psychological Review. 3 (6): 611–617. doi:10.1037/h0072918.
- Stratton, George M. (1897). "Vision without inversion of the retinal image". Psychological Review. 4 (4): 341–360, 463–481. doi:10.1037/h0075482.
- Hartmann, G.M. (1935). Gestalt Psychology. New York: The Ronald Press.
- Urbantschitsch, V. (1888). "Über den Einfluss einer Sinneserregung auf die übrigen Sinnesempfindungen". Pflügers Archiv. 42: 154–182. doi:10.1007/bf01669354. S2CID 42136599.
- Kravkov, S.V. (1930). "Über die Abhängigkeit der Sehschärfe vom Schallreiz". Arch. Ophthalmol. 124 (2): 334–338. doi:10.1007/bf01853661. S2CID 30040170.
- Hartmann, G.W. (1933). "Changes in Visual Acuity through Simultaneous Stimulation of Other Sense Organs". J. Exp. Psychol. 16 (3): 393–407. doi:10.1037/h0074549.
- London, I.D. (1954). "Research of sensory interaction in the Soviet Union". Psychol. Bull. 51 (6): 531–568. doi:10.1037/h0056730. PMID 13215683.
- Gonzalo, J. (1945, 1950, 1952, 2010, 2021). Dinámica Cerebral, Open Access. Edición facsímil 2010 del Vol. 1 1945, Vol. 2 1950 (Madrid: Inst. S. Ramón y Cajal, CSIC), Suplemento I 1952 (Trab. Inst. Cajal Invest. Biol.) y 1ª ed. Suplemento II 2010. Red Temática en Tecnologías de Computación Artificial/Natural (RTNAC) y Universidad de Santiago de Compostela (USC). ISBN 978-84-9887-458-7. English translation of Vol. 1 1945 (2021) Open Access. English translation of the article of 1952 (2015) Open Access.
- Gonzalo, J. (1952). "Las funciones cerebrales humanas según nuevos datos y bases fisiológicas. Una introducción a los estudios de Dinámica Cerebral". Trabajos del Inst. Cajal de Investigaciones Biológicas, XLIV: pp. 95–157. Complete English translation, Open Access.
- Witten, IB.; Knudsen, EI. (Nov 2005). "Why seeing is believing: merging auditory and visual worlds". Neuron. 48 (3): 489–96. doi:10.1016/j.neuron.2005.10.020. PMID 16269365. S2CID 17244783.
- Shams, L.; Beierholm, UR. (Sep 2010). "Causal inference in perception". Trends Cogn Sci. 14 (9): 425–32. doi:10.1016/j.tics.2010.07.001. PMID 20705502. S2CID 7750709.
- Gori, M.; Del Viva, M.; Sandini, G.; Burr, DC. (May 2008). "Young children do not integrate visual and haptic form information" (PDF). Curr Biol. 18 (9): 694–8. doi:10.1016/j.cub.2008.04.036. PMID 18450446. S2CID 13899031.
- Gori, M.; Sandini, G.; Burr, D. (2012). "Development of visuo-auditory integration in space and time". Front Integr Neurosci. 6: 77. doi:10.3389/fnint.2012.00077. PMC 3443931. PMID 23060759.
- Welch RB, Warren DH (November 1980). "Immediate perceptual response to intersensory discrepancy". Psychol Bull. 88 (3): 638–67. doi:10.1037/0033-2909.88.3.638. PMID 7003641.
- Lederman, Susan J.; Klatzky, Roberta L. (2004). "Multisensory Texture Perception". In Calvert, Gemma A.; Spence, Charles; Stein, Barry E. (eds.). The Handbook of Multisensory Processing. Cambridge, MA: MIT Press. pp. 107–122. ISBN 978-0-262-03321-3.
- Alais D, Burr D (February 2004). "The ventriloquist effect results from near-optimal bimodal integration". Curr. Biol. 14 (3): 257–62. CiteSeerX 10.1.1.220.4159. doi:10.1016/j.cub.2004.01.029. PMID 14761661. S2CID 3125842.
- Kuang, S.; Zhang, T. (2014). "Smelling directions: Olfaction modulates ambiguous visual motion perception". Scientific Reports. 4: 5796. Bibcode:2014NatSR...4E5796K. doi:10.1038/srep05796. PMC 4107342. PMID 25052162.
- Deneve S, Pouget A (2004). "Bayesian multisensory integration and cross-modal spatial links" (PDF). J. Physiol. Paris. 98 (1–3): 249–58. CiteSeerX 10.1.1.133.7694. doi:10.1016/j.jphysparis.2004.03.011. PMID 15477036. S2CID 9831111.
- Pouget A, Deneve S, Duhamel JR (September 2002). "A computational perspective on the neural basis of multisensory spatial representations". Nature Reviews Neuroscience. 3 (9): 741–7. doi:10.1038/nrn914. PMID 12209122. S2CID 1035721.
- Vilares, I.; Kording, K. (Apr 2011). "Bayesian models: the structure of the world, uncertainty, behavior, and the brain". Annals of the New York Academy of Sciences. 1224 (1): 22–39. Bibcode:2011NYASA1224...22V. doi:10.1111/j.1749-6632.2011.05965.x. PMC 3079291. PMID 21486294.
- Beierholm, UR.; Quartz, SR.; Shams, L. (2009). "Bayesian priors are encoded independently from likelihoods in human multisensory perception". J Vis. 9 (5): 23.1–9. doi:10.1167/9.5.23. PMID 19757901.
- Stein, Barry; Meredith, M. Alex (1993). The merging of the senses. Cambridge, Mass: MIT Press. ISBN 978-0-262-19331-3. OCLC 25869284.
- Meredith, MA.; Stein, BE. (Feb 1986). "Spatial factors determine the activity of multisensory neurons in cat superior colliculus". Brain Res. 365 (2): 350–4. doi:10.1016/0006-8993(86)91648-3. PMID 3947999. S2CID 12807282.
- King AJ, Palmer AR (1985). "Integration of visual and auditory information in bimodal neurones in the guinea-pig superior colliculus". Exp Brain Res. 60 (3): 492–500. doi:10.1007/bf00236934. PMID 4076371. S2CID 25796198.
- Meredith, MA.; Nemitz, JW.; Stein, BE. (Oct 1987). "Determinants of multisensory integration in superior colliculus neurons. I. Temporal factors". J Neurosci. 7 (10): 3215–29. doi:10.1523/JNEUROSCI.07-10-03215.1987. PMC 6569162. PMID 3668625.
- Meredith MA, Stein BE (July 1983). "Interactions among converging sensory inputs in the superior colliculus". Science. 221 (4608): 389–91. Bibcode:1983Sci...221..389M. doi:10.1126/science.6867718. PMID 6867718.
- Meredith, MA.; Stein, BE. (Sep 1986). "Visual, auditory, and somatosensory convergence on cells in superior colliculus results in multisensory integration". J Neurophysiol. 56 (3): 640–62. doi:10.1152/jn.1986.56.3.640. PMID 3537225.
- Cervantes Constantino, F.; Sánchez-Costa, T.; Cipriani, G.A.; Carboni, A. (2023). "Visuospatial attention revamps cortical processing of sound amid audiovisual uncertainty". Psychophysiology: e14329. doi:10.1111/psyp.14329. PMID 37166096. S2CID 258617930.
- Hershenson M (March 1962). "Reaction time as a measure of intersensory facilitation". J Exp Psychol. 63 (3): 289–93. doi:10.1037/h0039516. PMID 13906889.
- Hughes, HC.; Reuter-Lorenz, PA.; Nozawa, G.; Fendrich, R. (Feb 1994). "Visual-auditory interactions in sensorimotor processing: saccades versus manual responses". J Exp Psychol Hum Percept Perform. 20 (1): 131–53. doi:10.1037/0096-1523.20.1.131. PMID 8133219.
- Wallace, Mark T. (2004). "The development of multisensory processes". Cognitive Processing. 5 (2): 69–83. doi:10.1007/s10339-004-0017-z. ISSN 1612-4782. S2CID 16710851.
- Ridgway N, Milders M, Sahraie A (May 2008). "Redundant target effect and the processing of colour and luminance". Exp Brain Res. 187 (1): 153–60. doi:10.1007/s00221-008-1293-0. PMID 18264703. S2CID 23092762.
- Forster B, Cavina-Pratesi C, Aglioti SM, Berlucchi G (April 2002). "Redundant target effect and intersensory facilitation from visual-tactile interactions in simple reaction time". Exp Brain Res. 143 (4): 480–7. doi:10.1007/s00221-002-1017-9. PMID 11914794. S2CID 115844.
- McGurk H, MacDonald J (1976). "Hearing lips and seeing voices". Nature. 264 (5588): 746–8. Bibcode:1976Natur.264..746M. doi:10.1038/264746a0. PMID 1012311. S2CID 4171157.
- Nath, AR.; Beauchamp, MS. (Jan 2012). "A neural basis for interindividual differences in the McGurk effect, a multisensory speech illusion". NeuroImage. 59 (1): 781–7. doi:10.1016/j.neuroimage.2011.07.024. PMC 3196040. PMID 21787869.
- Hairston WD, Wallace MT, Vaughan JW, Stein BE, Norris JL, Schirillo JA (January 2003). "Visual localization ability influences cross-modal bias". J Cogn Neurosci. 15 (1): 20–9. doi:10.1162/089892903321107792. PMID 12590840. S2CID 13636325.
- Shams L, Kamitani Y, Shimojo S (December 2000). "Illusions. What you see is what you hear". Nature. 408 (6814): 788. Bibcode:2000Natur.408..788S. doi:10.1038/35048669. PMID 11130706. S2CID 205012107.
- Watkins S, Shams L, Josephs O, Rees G (August 2007). "Activity in human V1 follows multisensory perception". NeuroImage. 37 (2): 572–8. doi:10.1016/j.neuroimage.2007.05.027. PMID 17604652. S2CID 17477883.
- Shams L, Iwaki S, Chawla A, Bhattacharya J (April 2005). "Early modulation of visual cortex by sound: an MEG study". Neurosci. Lett. 378 (2): 76–81. doi:10.1016/j.neulet.2004.12.035. PMID 15774261. S2CID 4675944.
- Andersen TS, Tiippana K, Sams M (November 2004). "Factors influencing audiovisual fission and fusion illusions". Brain Res Cogn Brain Res. 21 (3): 301–308. doi:10.1016/j.cogbrainres.2004.06.004. PMID 15511646.
- Botvinick M, Cohen J (February 1998). "Rubber hands 'feel' touch that eyes see" (PDF). Nature. 391 (6669): 756. Bibcode:1998Natur.391..756B. doi:10.1038/35784. PMID 9486643. S2CID 205024516.
- Ehrsson HH, Holmes NP, Passingham RE (November 2005). "Touching a rubber hand: feeling of body ownership is associated with activity in multisensory brain areas". J. Neurosci. 25 (45): 10564–73. doi:10.1523/JNEUROSCI.0800-05.2005. PMC 1395356. PMID 16280594.
- Samad M, Chung A, Shams L (February 2015). "Perception of Body Ownership is Driven by Bayesian Sensory Inference". PLOS ONE. 10 (2): e0117178. Bibcode:2015PLoSO..1017178S. doi:10.1371/journal.pone.0117178. PMC 4320053. PMID 25658822.
- Holmes NP, Crozier G, Spence C (June 2004). "When mirrors lie: "visual capture" of arm position impairs reaching performance" (PDF). Cogn Affect Behav Neurosci. 4 (2): 193–200. doi:10.3758/CABN.4.2.193. PMC 1314973. PMID 15460925.
- J. Tastevin (Feb 1937). "En partant de l'expérience d'Aristote: Les déplacements artificiels des parties du corps ne sont pas suivis par le sentiment de ces parties ni par les sensations qu'on peut y produire" [Starting from Aristotle's experiment: The artificial displacements of parts of the body are not followed by feeling in these parts or by the sensations which can be produced there]. L'Encephale (in French). 32 (2): 57–84. (English abstract)
- J. Tastevin (Mar 1937). "En partant de l'expérience d'Aristote". L'Encephale (in French). 32 (3): 140–158.
- Bergman, Ronald A.; Afifi, Adel K. (2005). Functional neuroanatomy: text and atlas. New York: McGraw-Hill. ISBN 978-0-07-140812-7. OCLC 475017241.
- Giard MH, Peronnet F (September 1999). "Auditory-visual integration during multimodal object recognition in humans: a behavioral and electrophysiological study". J Cogn Neurosci. 11 (5): 473–90. doi:10.1162/089892999563544. PMID 10511637. S2CID 5735865.
- Miller LM, D'Esposito M (June 2005). "Perceptual fusion and stimulus coincidence in the cross-modal integration of speech". J. Neurosci. 25 (25): 5884–93. doi:10.1523/JNEUROSCI.0896-05.2005. PMC 6724802. PMID 15976077.
- Graziano MS, Gross CG (1993). "A bimodal map of space: somatosensory receptive fields in the macaque putamen with corresponding visual receptive fields" (PDF). Exp Brain Res. 97 (1): 96–109. doi:10.1007/BF00228820. PMID 8131835. S2CID 9387557.
- Gentile, G.; Petkova, VI.; Ehrsson, HH. (Feb 2011). "Integration of visual and tactile signals from the hand in the human brain: an FMRI study". J Neurophysiol. 105 (2): 910–22. doi:10.1152/jn.00840.2010. PMC 3059180. PMID 21148091.
- Thesen, T., Vibell, J., Calvert, G.A., & Osterbauer, R. (2004). Neuroimaging of multisensory processing in vision, audition, touch and olfaction. Cognitive Processing, 5, 84-93.
- Wallace MT, Ramachandran R, Stein BE (February 2004). "A revised view of sensory cortical parcellation". Proc. Natl. Acad. Sci. U.S.A. 101 (7): 2167–72. Bibcode:2004PNAS..101.2167W. doi:10.1073/pnas.0305697101. PMC 357070. PMID 14766982.
- Clavagnier S, Falchier A, Kennedy H (June 2004). "Long-distance feedback projections to area V1: implications for multisensory integration, spatial awareness, and visual consciousness" (PDF). Cogn Affect Behav Neurosci. 4 (2): 117–26. doi:10.3758/CABN.4.2.117. PMID 15460918. S2CID 6907281.
- Foxe JJ, Simpson GV (January 2002). "Flow of activation from V1 to frontal cortex in humans. A framework for defining "early" visual processing". Exp Brain Res. 142 (1): 139–50. doi:10.1007/s00221-001-0906-7. PMID 11797091. S2CID 25506401.
- Macaluso E, Frith CD, Driver J (August 2000). "Modulation of human visual cortex by crossmodal spatial attention". Science. 289 (5482): 1206–8. Bibcode:2000Sci...289.1206M. CiteSeerX 10.1.1.420.5403. doi:10.1126/science.289.5482.1206. PMID 10947990.
- Sadato N, Yamada H, Okada T, et al. (December 2004). "Age-dependent plasticity in the superior temporal sulcus in deaf humans: a functional MRI study". BMC Neurosci. 5: 56. doi:10.1186/1471-2202-5-56. PMC 539237. PMID 15588277.
- Bernstein LE, Auer ET, Moore JK, Ponton CW, Don M, Singh M (March 2002). "Visual speech perception without primary auditory cortex activation". NeuroReport. 13 (3): 311–5. doi:10.1097/00001756-200203040-00013. PMID 11930129. S2CID 44484836.
- Jones EG, Powell TP (1970). "An anatomical study of converging sensory pathways within the cerebral cortex of the monkey". Brain. 93 (4): 793–820. doi:10.1093/brain/93.4.793. PMID 4992433.
- Grefkes, C.; Fink, GR. (Jul 2005). "The functional organization of the intraparietal sulcus in humans and monkeys". J Anat. 207 (1): 3–17. doi:10.1111/j.1469-7580.2005.00426.x. PMC 1571496. PMID 16011542.
- Murata, A.; Fadiga, L.; Fogassi, L.; Gallese, V.; Raos, V.; Rizzolatti, G. (Oct 1997). "Object representation in the ventral premotor cortex (area F5) of the monkey". J Neurophysiol. 78 (4): 2226–30. doi:10.1152/jn.1997.78.4.2226. PMID 9325390.
- Smiley, JF.; Falchier, A. (Dec 2009). "Multisensory connections of monkey auditory cerebral cortex". Hear Res. 258 (1–2): 37–46. doi:10.1016/j.heares.2009.06.019. PMC 2788085. PMID 19619628.
- Sharma, J.; Dragoi, V.; Tenenbaum, JB.; Miller, EK.; Sur, M. (Jun 2003). "V1 neurons signal acquisition of an internal representation of stimulus location". Science. 300 (5626): 1758–63. Bibcode:2003Sci...300.1758S. doi:10.1126/science.1081721. PMID 12805552. S2CID 14716502.
- Lacey, S.; Tal, N.; Amedi, A.; Sathian, K. (May 2009). "A putative model of multisensory object representation". Brain Topogr. 21 (3–4): 269–74. doi:10.1007/s10548-009-0087-4. PMC 3156680. PMID 19330441.
- Neal, JW.; Pearson, RC.; Powell, TP. (Jul 1990). "The ipsilateral cortico-cortical connections of area 7b, PF, in the parietal and temporal lobes of the monkey". Brain Res. 524 (1): 119–32. doi:10.1016/0006-8993(90)90500-B. PMID 1698108. S2CID 24535669.
- Eickhoff, SB.; Schleicher, A.; Zilles, K.; Amunts, K. (Feb 2006). "The human parietal operculum. I. Cytoarchitectonic mapping of subdivisions". Cereb Cortex. 16 (2): 254–67. doi:10.1093/cercor/bhi105. PMID 15888607.
- Petitto LA, Zatorre RJ, Gauna K, Nikelski EJ, Dostie D, Evans AC (December 2000). "Speech-like cerebral activity in profoundly deaf people processing signed languages: implications for the neural basis of human language". Proc. Natl. Acad. Sci. U.S.A. 97 (25): 13961–6. doi:10.1073/pnas.97.25.13961. PMC 17683. PMID 11106400.
- Meredith, MA.; Clemo, HR. (Nov 1989). "Auditory cortical projection from the anterior ectosylvian sulcus (Field AES) to the superior colliculus in the cat: an anatomical and electrophysiological study". J Comp Neurol. 289 (4): 687–707. doi:10.1002/cne.902890412. PMID 2592605. S2CID 221577963.
- Jiang, W.; Wallace, MT.; Jiang, H.; Vaughan, JW.; Stein, BE. (Feb 2001). "Two cortical areas mediate multisensory integration in superior colliculus neurons". J Neurophysiol. 85 (2): 506–22. doi:10.1152/jn.2001.85.2.506. PMID 11160489. S2CID 2499047.
- Wallace, MT.; Carriere, BN.; Perrault, TJ.; Vaughan, JW.; Stein, BE. (Nov 2006). "The development of cortical multisensory integration". Journal of Neuroscience. 26 (46): 11844–9. doi:10.1523/JNEUROSCI.3295-06.2006. PMC 6674880. PMID 17108157.
- Jiang W, Stein BE (October 2003). "Cortex controls multisensory depression in superior colliculus". J. Neurophysiol. 90 (4): 2123–35. doi:10.1152/jn.00369.2003. PMID 14534263.
- Wallace MT, Meredith MA, Stein BE (June 1993). "Converging influences from visual, auditory, and somatosensory cortices onto output neurons of the superior colliculus". J. Neurophysiol. 69 (6): 1797–809. doi:10.1152/jn.1993.69.6.1797. PMID 8350124.
- Stein, Barry E.; London, Nancy; Wilkinson, Lee K.; Price, Donald D. (1996). "Enhancement of Perceived Visual Intensity by Auditory Stimuli: A Psychophysical Analysis". Journal of Cognitive Neuroscience. 8 (6): 497–506. doi:10.1162/jocn.1996.8.6.497. PMID 23961981. S2CID 43705477.
- Goodale MA, Milner AD (January 1992). "Separate visual pathways for perception and action" (PDF). Trends Neurosci. 15 (1): 20–5. CiteSeerX 10.1.1.207.6873. doi:10.1016/0166-2236(92)90344-8. PMID 1374953. S2CID 793980.
- Renier LA, Anurova I, De Volder AG, Carlson S, VanMeter J, Rauschecker JP (September 2009). "Multisensory integration of sounds and vibrotactile stimuli in processing streams for "what" and "where"". J. Neurosci. 29 (35): 10950–60. doi:10.1523/JNEUROSCI.0910-09.2009. PMC 3343457. PMID 19726653.
- Nardini, M; Bedford, R; Mareschal, D (Sep 28, 2010). "Fusion of visual cues is not mandatory in children". Proceedings of the National Academy of Sciences of the United States of America. 107 (39): 17041–6. Bibcode:2010PNAS..10717041N. doi:10.1073/pnas.1001699107. PMC 2947870. PMID 20837526.
- Gori, M; Del Viva, M; Sandini, G; Burr, DC (May 6, 2008). "Young children do not integrate visual and haptic form information" (PDF). Current Biology. 18 (9): 694–8. doi:10.1016/j.cub.2008.04.036. PMID 18450446. S2CID 13899031.
- Lewkowicz, D; Kraebel, K (2004). Gemma Calvert; Charles Spence; Barry E Stein (eds.). The value of multisensory redundancy in the development of intersensory perception. pp. 655–78. ISBN 9780262033213. OCLC 803222288.
{{cite book}}:|work=ignored (help) - Neil, PA; Chee-Ruiter, C; Scheier, C; Lewkowicz, DJ; Shimojo, S (Sep 2006). "Development of multisensory spatial integration and perception in humans". Developmental Science. 9 (5): 454–64. doi:10.1111/j.1467-7687.2006.00512.x. PMID 16911447. S2CID 25690976.
- Hillis, JM; Ernst, MO; Banks, MS; Landy, MS (Nov 22, 2002). "Combining sensory information: mandatory fusion within, but not between, senses". Science. 298 (5598): 1627–30. Bibcode:2002Sci...298.1627H. CiteSeerX 10.1.1.278.6134. doi:10.1126/science.1075396. PMID 12446912. S2CID 15607270.
- Ernst, MO (Jun 24, 2008). "Multisensory integration: a late bloomer". Current Biology. 18 (12): R519–21. doi:10.1016/j.cub.2008.05.002. PMID 18579094. S2CID 130911.
- Stein, BE; Meredith, MA; Wallace, MT (1993). The visually responsive neuron and beyond: multisensory integration in cat and monkey. Progress in Brain Research. Vol. 95. pp. 79–90. doi:10.1016/s0079-6123(08)60359-3. ISBN 9780444894922. PMID 8493355.
- Stein, BE; Labos, E; Kruger, L (Jul 1973). "Sequence of changes in properties of neurons of superior colliculus of the kitten during maturation". Journal of Neurophysiology. 36 (4): 667–79. doi:10.1152/jn.1973.36.4.667. PMID 4713313.
- Wallace, MT; Stein, BE (Nov 15, 2001). "Sensory and multisensory responses in the newborn monkey superior colliculus". The Journal of Neuroscience. 21 (22): 8886–94. doi:10.1523/JNEUROSCI.21-22-08886.2001. PMC 6762279. PMID 11698600.
- Tootell, RB; Hadjikhani, NK; Vanduffel, W; Liu, AK; Mendola, JD; Sereno, MI; Dale, AM (Feb 3, 1998). "Functional analysis of primary visual cortex (V1) in humans". Proceedings of the National Academy of Sciences of the United States of America. 95 (3): 811–7. Bibcode:1998PNAS...95..811T. doi:10.1073/pnas.95.3.811. PMC 33802. PMID 9448245.
- Gori, M; Giuliana, L; Sandini, G; Burr, D (Nov 2012). "Visual size perception and haptic calibration during development". Developmental Science. 15 (6): 854–62. doi:10.1111/j.1467-7687.2012.2012.01183.x. PMID 23106739.
- Granrud, CE; Schmechel, TT (Nov 2006). "Development of size constancy in children: a test of the proximal mode sensitivity hypothesis". Perception & Psychophysics. 68 (8): 1372–81. doi:10.3758/bf03193736. PMID 17378423.
- Gepshtein, S; Burge, J; Ernst, MO; Banks, MS (Dec 28, 2005). "The combination of vision and touch depends on spatial proximity". Journal of Vision. 5 (11): 1013–23. doi:10.1167/5.11.7. PMC 2632311. PMID 16441199.
- Helbig, HB; Ernst, MO (Jun 2007). "Optimal integration of shape information from vision and touch". Experimental Brain Research. 179 (4): 595–606. doi:10.1007/s00221-006-0814-y. PMID 17225091. S2CID 12049308.
- Helbig, HB; Ernst, MO (2007). "Knowledge about a common source can promote visual- haptic integration". Perception. 36 (10): 1523–33. doi:10.1068/p5851. PMID 18265835. S2CID 14884284.
- Kail, RV; Ferrer, E (Nov–Dec 2007). "Processing speed in childhood and adolescence: longitudinal models for examining developmental change". Child Development. 78 (6): 1760–70. doi:10.1111/j.1467-8624.2007.01088.x. PMID 17988319.
- Kail, R (May 1991). "Developmental change in speed of processing during childhood and adolescence". Psychological Bulletin. 109 (3): 490–501. doi:10.1037/0033-2909.109.3.490. PMID 2062981.
- Ballard, DH; Hayhoe, MM; Pook, PK; Rao, RP (Dec 1997). "Deictic codes for the embodiment of cognition". The Behavioral and Brain Sciences. 20 (4): 723–42, discussion 743–67. CiteSeerX 10.1.1.49.3813. doi:10.1017/s0140525x97001611. PMID 10097009. S2CID 1961389.
- Daee, Pedram; Mirian, Maryam S.; Ahmadabadi, Majid Nili (2014). "Reward Maximization Justifies the Transition from Sensory Selection at Childhood to Sensory Integration at Adulthood". PLOS ONE. 9 (7): e103143. Bibcode:2014PLoSO...9j3143D. doi:10.1371/journal.pone.0103143. PMC 4110011. PMID 25058591.
- Rincon-Gonzalez L, WarrenJ P (2011). "Haptic interaction of touch and proprioception: implications for neuroprosthetics". IEEE Trans. Neural Syst. Rehabil. Eng. 19 (5): 490–500. doi:10.1109/tnsre.2011.2166808. PMID 21984518. S2CID 20575638.
- Jiang, Huai; Stein, Barry E.; McHaffie, John G. (2015-05-29). "Multisensory training reverses midbrain lesion-induced changes and ameliorates haemianopia". Nature Communications. 6: 7263. Bibcode:2015NatCo...6.7263J. doi:10.1038/ncomms8263. ISSN 2041-1723. PMC 6193257. PMID 26021613.
Further reading
- Kujala, T.; Alho, K.; Huotilainen, M.; Ilmoniemi, RJ.; Lehtokoski, A.; Leinonen, A.; Rinne, T.; et, al. (Mar 1997). "Electrophysiological evidence for cross-modal plasticity in humans with early- and late-onset blindness". Psychophysiology. 34 (2): 213–6. doi:10.1111/j.1469-8986.1997.tb02134.x. PMID 9090272.
- Pascual-Leone, A.; Theoret, H.; et, al. (2006). Morton A Heller; Soledad Ballesteros (eds.). The Role of Visual Cortex in Tactile Processing: A Metamodal Brain. ISBN 9780805847260. OCLC 6124743.
{{cite book}}:|work=ignored (help) - Wallace, Mark; Murray, Micah Middelmann (2012). The Neural Bases of Multisensory Processes (Frontiers in Neuroscience). Boca Raton: CRC Press. ISBN 978-1-4398-1217-4. OCLC 707710852.