Min System
The Min System is a mechanism composed of three proteins MinC, MinD, and MinE used by E. coli as a means of properly localizing the septum prior to cell division. Each component participates in generating a dynamic oscillation of FtsZ protein inhibition between the two bacterial poles to precisely specify the mid-zone of the cell, allowing the cell to accurately divide in two. This system is known to function in conjunction with a second negative regulatory system, the nucleoid occlusion system (NO), to ensure proper spatial and temporal regulation of chromosomal segregation and division.
History
The initial discovery of this family of proteins is attributed to Adler et al. (1967). First identified as E. coli mutants that could not produce a properly localized septum, resulting in the generation of minicells[1][2] due to mislocalized cell division occurring near the bacterial poles. This caused miniature vesicles to pinch off, void of essential molecular constituents permitting it to exist as a viable bacterial cell. Minicells are achromosomal cells that are products of aberrant cell division, and contain RNA and protein, but little or no chromosomal DNA. This finding led to the identification of three interacting proteins involved in a dynamic system of localizing the mid-zone of the cell for properly controlled cell division.
Function
The Min proteins prevent the FtsZ ring from being placed anywhere but near the mid cell and are hypothesized to be involved in a spatial regulatory mechanism that links size increases prior to cell division to FtsZ polymerization in the middle of the cell.
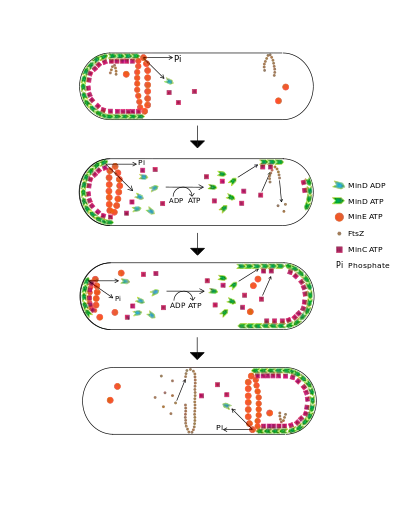
Centering the Z-Ring
One model of Z-ring formation permits its formation only after a certain spatial signal that tells the cell that it is big enough to divide.[3] The MinCDE system prevents FtsZ polymerization near certain parts of the plasma membrane. MinD localizes to the membrane only at cell poles and contains an ATPase and an ATP-binding domain. MinD is only able to bind to the membrane when in its ATP-bound conformation. Once anchored, the protein polymerizes, resulting in clusters of MinD. These clusters bind and then activate another protein called MinC, which has activity only when bound by MinD.[4] MinC serves as a FtsZ inhibitor that prevents FtsZ polymerization. The high concentration of a FtsZ polymerization inhibitor at the poles prevents FtsZ from initiating division at anywhere but the mid-cell.[5]
MinE is involved in preventing the formation of MinCD complexes in the middle of the cell. MinE forms a ring near each cell pole. This ring is not like the Z-ring. Instead, it catalyzes the release of MinD from the membrane by activating MinD's ATPase. This hydrolyzes the MinD's bound ATP, preventing it from anchoring itself to the membrane.
MinE prevents the MinD/C complex from forming in the center but allows it to stay at the poles. Once the MinD/C complex is released, MinC becomes inactivated. This prevents MinC from deactivating FtsZ. As a consequence, this activity imparts regional specificity to Min localization.[6] Thus, FtsZ can form only in the center, where the concentration of the inhibitor MinC is minimal. Mutations that prevent the formation of MinE rings result in the MinCD zone extending well beyond the polar zones, preventing FtsZ to polymerize and to perform cell division.[7] MinD requires a nucleotide exchange step to re-bind to ATP so that it can re-associate with the membrane after MinE release. The time lapse results in a periodicity of Min association that may yield clues to a temporal signal linked to a spatial signal. In vivo observations show that the oscillation of Min proteins between cell poles occurs approximately every 50 seconds.[8] Oscillation of Min proteins, however, is not necessary for all bacterial cell division systems. Bacillus subtilis has been shown to have static concentrations of MinC and MinD at the cell poles.[9] This system still links cell size to the ability to form a septum via FtsZ and divide.
in vitro Reconstitution

The dynamic behavior of Min proteins has been reconstituted in vitro using artificial lipid bilayers,[10] with varying lipid composition[11] and different confinement geometry[12] as mimics for the cell membrane. The first pattern to be reconstituted were spiraling waves of MinD chased by MinE,[13] followed by the reconstitution of waves of all three proteins, MinD, MinE and MinC.[14] Importantly, MinD and MinE can self-organize into a wide variety of patterns depending on the reaction conditions.[15][16]
Additional study is required to elucidate the extent of temporal and spatial signaling permissible by this biological function. These in vitro systems offered unprecedented access to features such as residence times and molecular motility.
References
- De Boer PA, Crossley RE, Rothfield LI (1989). "A division inhibitor and a topological specificity factor coded for by the minicell locus determine proper placement of the division septum in E. coli". Cell. 56 (4): 641–649. doi:10.1016/0092-8674(89)90586-2. PMID 2645057. S2CID 7650379.
- Adler HI, Fisher WD, Cohen A, Hardigree AA; Fisher; Cohen; Hardigree (1967). "Miniature Escherichia coli Cells Deficient in DNA". PNAS. 57 (2): 321–326. Bibcode:1967PNAS...57..321A. doi:10.1073/pnas.57.2.321. PMC 335508. PMID 16591472.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Weart RB, Levin PA (2003). "Growth Rate-Dependent Regulation of Medial FtsZ Ring Formation". J Bacteriol. 185 (9): 2826–2834. doi:10.1128/JB.185.9.2826-2834.2003. PMC 154409. PMID 12700262.
- Hu Z, Gogol EP, Lutkenhaus J (2002). "Dynamic assembly of MinD on phospholipid vesicles regulated by ATP and MinE". Proc Natl Acad Sci USA. 99 (10): 6761–6766. Bibcode:2002PNAS...99.6761H. doi:10.1073/pnas.102059099. PMC 124476. PMID 11983867.
- Huang KC, Meir Y, Wingreen NS (2003). "Dynamic structures in Escherichia coli: Spontaneous formation of MinE rings and MinD polar zones". Proc Natl Acad Sci USA. 100 (22): 12724–12728. Bibcode:2003PNAS..10012724H. doi:10.1073/pnas.2135445100. PMC 240685. PMID 14569005.
- Hu Z, Saez C, Lutkenhaus J (2003). "Recruitment of MinC, an Inhibitor of Z-Ring Formation, to the Membrane in Escherichia coli: Role of MinD and MinE". J Bacteriol. 185 (1): 196–203. doi:10.1128/JB.185.1.196-203.2003. PMC 141945. PMID 12486056.
- Hu Z, Lutkenhaus J (2001). "Topological regulation of cell division in E. coli: spatiotemporal oscillation of MinD requires stimulation of its ATPase by MinE and phospholipid". Mol Cell. 7 (6): 1337–1343. doi:10.1016/S1097-2765(01)00273-8. PMID 11430835.
- Dajkovic A, Lutkenhaus J (2006). "Z Ring as Executor of Bacterial Cell Division". J Mol Micro Bio. 11 (3–5): 140–151. doi:10.1159/000094050. PMID 16983191. S2CID 10043376.
- Marston AL, Thomaides HB, Edwards DH, Sharpe ME, Errington J (1998). "Polar localization of the MinD protein of Bacillus subtilis and its role in selection of the mid-cell division site". Genes Dev. 12 (21): 3419–3430. doi:10.1101/gad.12.21.3419. PMC 317235. PMID 9808628.
- Loose, Martin; Fischer-Friedrich, Elisabeth; Ries, Jonas; Kruse, Karsten; Schwille, Petra (2008-05-09). "Spatial regulators for bacterial cell division self-organize into surface waves in vitro". Science. 320 (5877): 789–792. Bibcode:2008Sci...320..789L. doi:10.1126/science.1154413. ISSN 1095-9203. PMID 18467587. S2CID 27134918.
- Vecchiarelli, Anthony G.; Li, Min; Mizuuchi, Michiyo; Mizuuchi, Kiyoshi (2014-08-01). "Differential affinities of MinD and MinE to anionic phospholipid influence Min patterning dynamics in vitro". Molecular Microbiology. 93 (3): 453–463. doi:10.1111/mmi.12669. ISSN 1365-2958. PMC 4116444. PMID 24930948.
- Zieske, Katja; Schwille, Petra (2014-01-01). "Reconstitution of self-organizing protein gradients as spatial cues in cell-free systems". eLife. 3. doi:10.7554/eLife.03949. ISSN 2050-084X. PMC 4215534. PMID 25271375.
- Loose, Martin; Fischer-Friedrich, Elisabeth; Ries, Jonas; Kruse, Karsten; Schwille, Petra (2008-05-09). "Spatial regulators for bacterial cell division self-organize into surface waves in vitro". Science. 320 (5877): 789–792. Bibcode:2008Sci...320..789L. doi:10.1126/science.1154413. ISSN 1095-9203. PMID 18467587. S2CID 27134918.
- Loose, Martin; Fischer-Friedrich, Elisabeth; Herold, Christoph; Kruse, Karsten; Schwille, Petra (2011-05-09). "Min protein patterns emerge from rapid rebinding and membrane interaction of MinE". Nat Struct Mol Biol. 18 (5): 577–83. doi:10.1038/nsmb.2037. PMID 21516096. S2CID 30014235.
- Ivanov, V.; Mizuuchi, K. (2010-03-08). "Multiple modes of interconverting dynamic pattern formation by bacterial cell division proteins". Proceedings of the National Academy of Sciences. 107 (18): 8071–8078. doi:10.1073/pnas.0911036107. ISSN 0027-8424. PMC 2889524. PMID 20212106.
- Vecchiarelli, Anthony G.; Li, Min; Mizuuchi, Michiyo; Hwang, Ling Chin; Seol, Yeonee; Neuman, Keir C.; Mizuuchi, Kiyoshi (2016-03-15). "Membrane-bound MinDE complex acts as a toggle switch that drives Min oscillation coupled to cytoplasmic depletion of MinD". Proceedings of the National Academy of Sciences of the United States of America. 113 (11): E1479–1488. Bibcode:2016PNAS..113E1479V. doi:10.1073/pnas.1600644113. ISSN 1091-6490. PMC 4801307. PMID 26884160.