Mesozoic–Cenozoic radiation
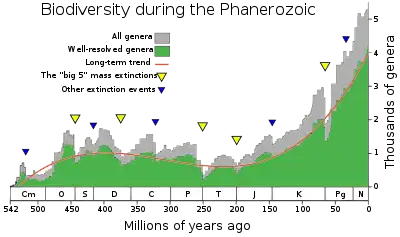
The Mesozoic–Cenozoic Radiation is the third major extended increase of biodiversity in the Phanerozoic,[1] after the Cambrian Explosion and the Great Ordovician Biodiversification Event, which appeared to exceeded the equilibrium reached after the Ordovician radiation. Made known by its identification in marine invertebrates, this evolutionary radiation began in the Mesozoic, after the Permian extinctions, and continues to this date. This spectacular radiation affected both terrestrial and marine flora and fauna,[2] during which the “modern” fauna came to replace much of the Paleozoic fauna.[1] Notably, this radiation event was marked by the rise of angiosperms during the mid-Cretaceous,[3] and the K-Pg extinction, which initiated the rapid increase in mammalian biodiversity.[4][5]

Causes and significance
The exact causes of this extended increase in biodiversity are still being debated, however, the Mesozoic-Cenozoic radiation has often been related to large-scale paleogeographical changes.[6][2][7] The fragmentation of the supercontinent Pangaea has been related to an increase in both marine and terrestrial biodiversity.[8][9][10] The link between the fragmentation of supercontinents and biodiversity was first proposed by Valentine and Moores in 1972. They hypothesized that the isolation of terrestrial environments and the partitioning of oceanic water masses, as a result of the breaking up of Pangaea, resulted in an increase in allopatric speciation, which led to an increased biodiversity.[11] These smaller landmasses, while individually being less diverse than a supercontinent, contain a high degree of endemic species, resulting in an overall higher biodiversity than a single landmass of equivalent size.[8] It is therefore argued that, similarly to the Ordovician bio-diversification, the differentiation of biotas along environmental gradients caused by the fragmentation of a supercontinent, was a driving force behind the Mesozoic-Cenozoic radiation.[6][7]
Part of the dramatic increase in biodiversity during this time was caused by the evolutionary radiation of flowering plants, or angiosperms, during the mid-Cretaceous.[3] Characteristics of this clade associated with reproduction have served as a key innovation for an entire clade, and led to a burst of evolution after the origin of the flower.[12] These later diversified further and co-radiated with pollinating insects, increasing biodiversity.[13]
A third factor which played a role in the Mesozoic-Cenozoic radiation was the K-Pg extinction, which marked the end of the dinosaurs and, surprisingly, resulted in a massive increase in biodiversity of terrestrial tetrapods, which can almost entirely be attributed to the radiation of mammals. There are multiple things which could have caused this deviation from the equilibrium, one of which is that the before the K-Pg extinction an equilibrium was reached which limited biodiversity.[4][5] The extinction event reorganized the fundamental ecology, on which diversity is built and maintained. After these reorganized ecosystems stabilized a new, higher, equilibrium was reached, which was maintained during the Cenozoic.[14]
Pull of the recent
One effect which has to be taken into account when estimating past biodiversity levels is the pull of the recent, which describes a phenomenon in the fossil record which causes biodiversity estimates to be skewed towards modern taxa.[15][16] This bias towards recent taxa is caused by a better availability of more recent fossil records. In mammals it has also been argued that the complexity of teeth, allowing for precise taxonomic identification of fragmentary fossils, increases their perceived diversity when compared to other clades at the time.[4][5] The contribution of this effect to the apparent increase in biodiversity is still unclear and heavily debated.[17][5][6]
See also
- Evolutionary faunas, the third of which corresponds to the Mesozoic–Cenozoic radiation
References
- Sepkoski, J. John (8 February 2016). "A factor analytic description of the Phanerozoic marine fossil record". Paleobiology. 7 (1): 36–53. doi:10.1017/S0094837300003778. S2CID 133114885.
- Owen, A.W.; Crame, J.A. (2002). "Palaeobiogeography and the Ordovician and Mesozoic-Cenozoic biotic radiations". Geological Society, London. Special Publications. 194 (1): 1–11. Bibcode:2002GSLSP.194....1O. doi:10.1144/GSL.SP.2002.194.01.01. S2CID 130743665.
- Crame, J. Alistair (July 2001). "Taxonomic diversity gradients through geological time". Diversity & Distributions. 7 (4): 175–189. doi:10.1111/j.1472-4642.2001.00106.x. S2CID 86824779.
- Benson, Roger B.J.; Butler, Richard J.; Alroy, John; Mannion, Philip D.; Carrano, Matthew T.; Lloyd, Graeme T.; Barnosky, Anthony D. (25 January 2016). "Near-Stasis in the Long-Term Diversification of Mesozoic Tetrapods". PLOS Biology. 14 (1): e1002359. doi:10.1371/journal.pbio.1002359. PMC 4726655. PMID 26807777.
- Close, Roger A.; Benson, Roger B. J.; Alroy, John; Behrensmeyer, Anna K.; Benito, Juan; Carrano, Matthew T.; Cleary, Terri J.; Dunne, Emma M.; Mannion, Philip D.; Uhen, Mark D.; Butler, Richard J. (18 February 2019). "Diversity dynamics of Phanerozoic terrestrial tetrapods at the local-community scale" (PDF). Nature Ecology & Evolution. 3 (4): 590–597. doi:10.1038/s41559-019-0811-8. PMID 30778186. S2CID 66884562.
- Crame, J.A.; Rosen, B.R. (2002). "Cenozoic palaeogeography and the rise of modern biodiversity patterns". Geological Society, London. Special Publications. 194 (1): 153–168. Bibcode:2002GSLSP.194..153C. doi:10.1144/GSL.SP.2002.194.01.12. S2CID 128674404.
- Servais, Thomas; Harper, David A.T.; Munnecke, Axel; Owen, Alan W.; Sheehan, Peter M. (2009). "Understanding the Great Ordovician Biodiversification Event (GOBE): Influences of paleogeography, paleoclimate, or paleoecology". GSA Today. 19 (4): 4. doi:10.1130/GSATG37A.1.
- Vavrek, Matthew J. (September 2016). "The fragmentation of Pangaea and Mesozoic terrestrial vertebrate biodiversity". Biology Letters. 12 (9): 20160528. doi:10.1098/rsbl.2016.0528. PMC 5046932. PMID 27651536.
- Zaffos, Andrew; Finnegan, Seth; Peters, Shanan E. (30 May 2017). "Plate tectonic regulation of global marine animal diversity". Proceedings of the National Academy of Sciences of the United States of America. 114 (22): 5653–5658. Bibcode:2017PNAS..114.5653Z. doi:10.1073/pnas.1702297114. PMC 5465924. PMID 28507147.
- Cermeño, Pedro; García-Comas, Carmen; Pohl, Alexandre; Williams, Simon; Benton, Michael J.; Chaudhary, Chhaya; Le Gland, Guillaume; Müller, R. Dietmar; Ridgwell, Andy; Vallina, Sergio M. (13 July 2022). "Post-extinction recovery of the Phanerozoic oceans and biodiversity hotspots". Nature. 607 (7919): 507–511. doi:10.1038/s41586-022-04932-6. ISSN 1476-4687. Retrieved 1 October 2023.
- Valentine, James W.; Moores, Eldridge M. (1972). "Global Tectonics and the Fossil Record". The Journal of Geology. 80 (2): 167–184. Bibcode:1972JG.....80..167V. doi:10.1086/627723. ISSN 0022-1376. JSTOR 30059313. S2CID 129364140.
- Soltis, Pamela S.; Folk, Ryan A.; Soltis, Douglas E. (27 March 2019). "Darwin review: Angiosperm phylogeny and evolutionary radiations". Proceedings of the Royal Society B: Biological Sciences. 286 (1899): 20190099. doi:10.1098/rspb.2019.0099. PMC 6452062.
- Grimaldi, David (Spring 1999). "The Co-Radiations of Pollinating Insects and Angiosperms in the Cretaceous". Annals of the Missouri Botanical Garden. 86 (2): 373–406. doi:10.2307/2666181. JSTOR 2666181.
- Bambach, R.K.; Knoll, Andrew H.; Sepkoski, J.J. (14 May 2002). "Anatomical and ecological constraints on Phanerozoic animal diversity in the marine realm". Proceedings of the National Academy of Sciences of the United States of America. 99 (10): 6854–6859. Bibcode:2002PNAS...99.6854B. doi:10.1073/pnas.092150999. PMC 124493. PMID 12011444.
- Alroy, J.; Aberhan, M.; Bottjer, D.J.; Foote, M.; Fursich, F.T.; Harries, P.J.; et al. (4 July 2008). "Phanerozoic Trends in the Global Diversity of Marine Invertebrates". Science. 321 (5885): 97–100. Bibcode:2008Sci...321...97A. doi:10.1126/science.1156963. PMID 18599780. S2CID 35793274.
- Ivany, Linda C.; Czekanski-Moir, Jesse (25 March 2019). "Reined-in richness". Nature Ecology & Evolution. 3 (4): 520–521. doi:10.1038/s41559-019-0863-9. PMID 30911149.
- Alroy, J.; Marshall, C.R.; Bambach, R.K.; Bezusko, K.; Foote, M.; Fursich, F.T.; et al. (15 May 2001). "Effects of sampling standardization on estimates of Phanerozoic marine diversification". Proceedings of the National Academy of Sciences of the United States of America. 98 (11): 6261–6266. doi:10.1073/pnas.111144698. PMC 33456. PMID 11353852.