Kleptoplasty
Kleptoplasty or kleptoplastidy is a process in symbiotic relationships whereby plastids, notably chloroplasts from algae, are sequestered by the host. The word is derived from Kleptes (κλέπτης) which is Greek for thief. The alga is eaten normally and partially digested, leaving the plastid intact. The plastids are maintained within the host, temporarily continuing photosynthesis and benefiting the host.
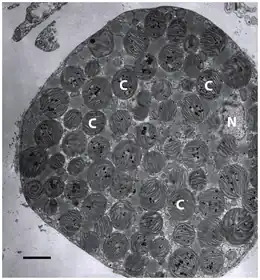
C = chloroplast,
N = cell nucleus.
Electron micrograph: scale bar is 3 µm.
Etymology
The word kleptoplasty is derived from Greek kleptes (κλέπτης), thief, and plastós (πλαστός), originally meaning formed or moulded, and used in biology to mean a plastid.[1]
Process
Kleptoplasty is a process in symbiotic relationships whereby plastids, notably chloroplasts from algae, are sequestered by the host. The alga is eaten normally and partially digested, leaving the plastid intact. The plastids are maintained within the host, temporarily continuing photosynthesis and benefiting the host.[1] The term was coined in 1990 to describe chloroplast symbiosis.[2][3]
Taxonomic range
Kleptoplasty occurs in two major clades of eukaryotes, namely micro-organisms of the SAR Supergroup, and some marine animals.
| Eukaryota |
| |||||||||||||||||||||||||||||||||
Foraminifera
Some species of the foraminiferan genera Bulimina, Elphidium, Haynesina, Nonion, Nonionella, Nonionellina, Reophax, and Stainforthia sequester diatom chloroplasts.[4]
Dinoflagellates
The stability of transient plastids varies considerably across plastid-retaining species. In the dinoflagellates Gymnodinium spp. and Pfisteria piscicida, kleptoplastids are photosynthetically active for only a few days, while kleptoplastids in Dinophysis spp. can be stable for 2 months.[1] In other dinoflagellates, kleptoplasty has been hypothesized to represent either a mechanism permitting functional flexibility, or perhaps an early evolutionary stage in the permanent acquisition of chloroplasts.[5]
Ciliates
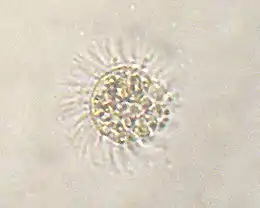
Mesodinium rubrum is a ciliate that steals chloroplasts from the cryptomonad Geminigera cryophila.[6] M. rubrum participates in additional endosymbiosis by transferring its plastids to its predators, the dinoflagellate planktons belonging to the genus Dinophysis.[7]
Karyoklepty is a related process in which the nucleus of the prey cell is kept by the host as well. This was first described in 2007 in M. rubrum.[8]
Rhabdocoel flatworms
Two species of rhabdocoel marine flatworms, Baicalellia solaris and Pogaina paranygulgus, make use of kleptoplasty. The group was previously classified as having algal endosymbionts, though it was already discovered that the endosymbionts did not contain nuclei.[9]
While consuming diatoms, B. solaris and P. paranygulus, in a process not yet discovered, extract plastids from their prey, incorporating them subepidermally, while separating and digesting the frustule and remainder of the diatom. In B. solaris the extracted plastids, or kleptoplasts, continue to exhibit functional photosynthesis for a short period of roughly 7 days. As the two groups are not sister taxa, and the trait is not shared among groups more closely related, there is evidence that kleptoplasty evolved independently within the two taxa.[10]
Sacoglossa
Sea slugs in the clade Sacoglossa practise kleptoplasty.[11] Several species of Sacoglossan sea slugs capture intact, functional chloroplasts from algal food sources, retaining them within specialized cells lining the mollusc's digestive diverticula. The longest known kleptoplastic association, which can last up to ten months, is found in Elysia chlorotica,[2] which acquires chloroplasts by eating the alga Vaucheria litorea, storing the chloroplasts in the cells that line its gut.[12] Juvenile sea slugs establish the kleptoplastic endosymbiosis when feeding on algal cells, sucking out the cell contents, and discarding everything except the chloroplasts. The chloroplasts are phagocytosed by digestive cells, filling extensively branched digestive tubules, providing their host with the products of photosynthesis.[13] It is not resolved, however, whether the stolen plastids actively secrete photosynthate or whether the slugs profit indirectly from slowly degrading kleptoplasts.[14]
Due to this unusual ability, the sacoglossans are sometimes referred to as "solar-powered sea slugs," though the actual benefit from photosynthesis on the survival of some of the species that have been analyzed seems to be marginal at best.[15] In fact, some species may even die in the presence of the carbon dioxide-fixing kleptoplasts as a result of elevated levels of reactive oxygen species.[16]
Changes in temperature have been shown to negatively affect kleptoplastic abilities in sacoglossans. Rates of photosynthetic efficiency as well as kleptoplast abundance have been shown to decrease in correlation to a decrease in temperature. The patterns and rate of these changes, however, varies between different species of sea slug.[17]
Nudibranchia
Some species of another group of sea slugs, nudibranchs such as Pteraeolidia ianthina, sequester whole living symbiotic zooxanthellae within their digestive diverticula, and thus are similarly "solar-powered".[18]
See also
References
- Minnhagen, Susanna; Carvalho, Wanderson F.; Salomon, Paulo S.; Janson, Sven (2008). "Chloroplast DNA content inDinophysis(Dinophyceae) from different cell cycle stages is consistent with kleptoplasty". Environmental Microbiology. Wiley. 10 (9): 2411–2417. doi:10.1111/j.1462-2920.2008.01666.x. PMID 18518896.
- S. K. Pierce; S. E. Massey; J. J. Hanten; N. E. Curtis (June 1, 2003). "Horizontal Transfer of Functional Nuclear Genes Between Multicellular Organisms". Biol. Bull. 204 (3): 237–240. doi:10.2307/1543594. JSTOR 1543594. PMID 12807700. S2CID 9671982. Retrieved 2008-11-24.
- Clark, K. B.; Jensen, K. R.; Strits, H. M. (1990). "Survey for Functional Kleptoplasty Among West Atlantic Ascoglossa (=Sacoglossa) (Mollusca: Opisthobranchia)". The Veliger. 33: 339–345. S2CID 87182226.
- Bernhard, Joan M.; Bowser, Samuel S. (1999). "Benthic foraminifera of dysoxic sediments: chloroplast sequestration and functional morphology". Earth-Science Reviews. 46 (1–4): 149–165. Bibcode:1999ESRv...46..149B. doi:10.1016/s0012-8252(99)00017-3.
- Gast RJ, Moran DM, Dennett MR, Caron DA (January 2007). "Kleptoplasty in an Antarctic dinoflagellate: caught in evolutionary transition?". Environ. Microbiol. 9 (1): 39–45. CiteSeerX 10.1.1.490.54. doi:10.1111/j.1462-2920.2006.01109.x. PMID 17227410.
- Johnson, Matthew D.; Oldach, David; Charles, F. Delwiche; Stoecker, Diane K. (Jan 2007). "Retention of transcriptionally active cryptophyte nuclei by the ciliate Myrionecta rubra". Nature. 445 (7126): 426–8. Bibcode:2007Natur.445..426J. doi:10.1038/nature05496. PMID 17251979. S2CID 4410812.
- Nishitani, G.; Nagai, S.; Baba, K.; Kiyokawa, S.; Kosaka, Y.; Miyamura, K.; Nishikawa, T.; Sakurada, K.; Shinada, A.; Kamiyama, T. (2010). "High-level congruence of Myrionecta rubra prey and Dinophysis species plastid identities as revealed by genetic analyses of isolates from Japanese coastal waters". Applied and Environmental Microbiology. 76 (9): 2791–2798. doi:10.1128/AEM.02566-09. PMC 2863437. PMID 20305031.
- Johnson, Matthew D.; Oldach, David; et al. (25 January 2007). "Retention of transcriptionally active cryptophyte nuclei by the ciliate Myrionecta rubra". Nature. 445 (7126): 426–428. Bibcode:2007Natur.445..426J. doi:10.1038/nature05496. PMID 17251979. S2CID 4410812. Retrieved 4 February 2015.
- E. Marcus, Turbellaria Brasileiros (9). Bol. Fac. Fil. Ci. Letras Univ. São Paulo 16, 5–215 (1951).
- Van Steenkiste, Niels W. L.; Stephenson, India; Herranz, María; Husnik, Filip; Keeling, Patrick J.; Leander, Brian S. (2019). "A new case of kleptoplasty in animals: Marine flatworms steal functional plastids from diatoms". Science Advances. 5 (7): eaaw4337. Bibcode:2019SciA....5.4337V. doi:10.1126/sciadv.aaw4337. PMC 6636991. PMID 31328166.
- Händeler, K.; Grzymbowski, Y. P.; Krug, P. J.; Wägele, H. (2009). "Functional chloroplasts in metazoan cells - a unique evolutionary strategy in animal life". Frontiers in Zoology. 6: 28. doi:10.1186/1742-9994-6-28. PMC 2790442. PMID 19951407.
- Catherine Brahic (24 November 2008). "Solar-powered sea slug harnesses stolen plant genes". New Scientist. Retrieved 24 November 2008.
- "SymBio: Introduction-Kleptoplasty". University of Maine. Archived from the original on 2008-12-02. Retrieved 2008-11-24.
- de Vries, Jan; Christa, Gregor; Gould, Sven B. (2014). "Plastid survival in the cytosol of animal cells". Trends in Plant Science. 19 (6): 347–350. doi:10.1016/j.tplants.2014.03.010. PMID 24767983.
- De Vries, Jan; Rauch, Cessa; Christa, Gregor; Gould, Sven B. (2014). "A sea slug's guide to plastid symbiosis". Acta Societatis Botanicorum Poloniae. 83 (4): 415–421. doi:10.5586/asbp.2014.042. ISSN 2083-9480.
- de Vries, J.; Woehle, C.; Christa, G.; Wagele, H.; Tielens, A. G. M.; Jahns, P.; Gould, S. B. (2015). "Comparison of sister species identifies factors underpinning plastid compatibility in green sea slugs". Proceedings of the Royal Society B: Biological Sciences. 282 (1802): 20142519. doi:10.1098/rspb.2014.2519. ISSN 0962-8452. PMC 4344150. PMID 25652835.
- Laetz, Elise; Wagele, Heike (2018). "How does temperature affect functional kleptoplasty? Comparing populations of the solar-powered sister-species Elysia timida Risso, 1818 and Elysa cornigera Nuttall, 1989 (Gastopoda: Sacoglossa)". Frontiers in Zoology. 15: 17. doi:10.1186/s12983-018-0264-y. PMC 5937827. PMID 29760759.
- Sutton & Hoegh-Guldberg, Host-Zooxanthella Interactions in Four Temperate Marine Symbioses; Assessment of Effect of Host Extract on Symbionts; The Biological bulletin, Marine Biological Laboratory (Woods Hole, Mass.). v. 178 (1990) p. 175
External links
- "Solar Powered Sea Slugs". ABC Science Online. June 2007. Retrieved 2008-11-24.