Juxtacrine signalling
In biology, juxtacrine signalling (or contact-dependent signalling) is a type of cell–cell or cell–extracellular matrix signalling in multicellular organisms that requires close contact. In this type of signalling, a ligand on one surface binds to a receptor on another adjacent surface. Hence, this stands in contrast to releasing a signaling molecule by diffusion into extracellular space, the use of long-range conduits like membrane nanotubes and cytonemes (akin to 'bridges') or the use of extracellular vesicles like exosomes or microvesicles (akin to 'boats'). There are three types of juxtacrine signaling:
- A membrane-bound ligand (protein, oligosaccharide, lipid) and a membrane protein of two adjacent cells interact.
- A communicating junction links the intracellular compartments of two adjacent cells, allowing transit of relatively small molecules.
- An extracellular matrix glycoprotein and a membrane protein interact.
Additionally, in unicellular organisms such as bacteria, juxtacrine signaling refers to interactions by membrane contact.
Juxtacrine signaling has been observed for some growth factors, cytokine and chemokine cellular signals, playing an important role in the immune response. It has a critical role in development, particularly of cardiac and neural function. Other types of cell signaling include paracrine signalling and autocrine signalling. Paracrine signaling occurs over short distances, while autocrine signaling involves a cell responding to its own paracrine factors.
The term "juxtacrine" was originally introduced by Anklesaria et al. (1990) to describe a possible way of signal transduction between TGF alpha and EGFR.[1]
Cell–cell signaling
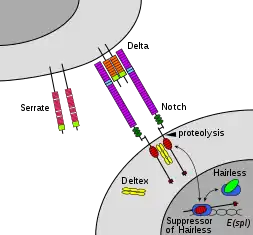
In this type of signaling, specific membrane-bound ligands bind to a cell’s membrane. A cell with the appropriate cell surface receptor or cell adhesion molecule can bind to it. An important example is the Notch signaling pathway, notably involved in neural development.[2] In the Notch signaling pathway for vertebrates and Drosophila, the receiving cell is told not to become neural through the binding of Delta and Notch. Within the eye of vertebrates, which cells become optic neurons and which become glial cells is regulated by Notch and its ligands.[3]
Some cells, like ephrin-Eph and cadherin, are only able to communicate through juxtacrine signaling. Eph ligands can only activate receptors when bound to a membrane.[4] This is because a high density of the Eph ligand is necessary for the receptor to bind to it.[5] Ephrin-Eph is used for axon guidance, angiogenesis, and epithelial and neuronal cell migration.[2][5]
Communicating junctions
Two adjacent cells can construct communicating conduits between their intracellular compartments: gap junctions in animals and plasmodesmas in plants.[2][6]
Gap junctions are made of connexins in vertebrates and innexins in invertebrates. Electrical synapses are electrically conductive gap junctions between neurons. Gap junctions are critical for cardiac myocytes; mice and humans deficient in a particular gap junction protein have severe heart development defects.[7]
Plasmodesmas in plants are cytoplasmic strands that pass through cell walls and facilitate connections with adjacent cells. Plasmodesmas are highly dynamic in both strucutural modifications and biogenesis. They are able to organize cells in domains, serving as basic developmental units for plants, as well as mediate the intracellular movement of a variety of proteins and nucleic acids.[8]
Cell–extracellular matrix signaling
The extracellular matrix is composed of glycoproteins (proteins and mucopolysaccharides (glycosaminoglycan)) produced by the organism's cells. They are secreted not only to build a supportive structure but also to provide critical information on the immediate environment to nearby cells. Indeed, the cells can themselves interact by contact with extracellular matrix molecules and as such, this can be considered an indirect cell / cell communication.[2] Cells use mainly the receptor integrin to interact with ECM proteins. Integrins are a family of receptor proteins that integrate the extracellular and intracellular structures, allowing them to perform together.[3] This signaling can influence the cell cycle and cellular differentiation by directing which cells live or die, which cells proliferate, or which cells are able to exit the cell cycle and differentiate.[9] Cellular differentiation involves a cell changing its phenotypical or functional type.
See also
- Cell adhesion, mechanical adhesion between cells and/or the extracellular matrix
- Role of cell adhesions in neural development
- Cell adhesion molecules
- Pannexin, vertebrate proteins used to form conduits between the intracellular and extracellular space
- Autocrine signalling
- Paracrine signalling
- Endocrine system
References
- Anklesaria, P; Teixidó, J; Laiho, M; Pierce, JH; Greenberger, JS; Massagué, J (May 1990). "Cell-cell adhesion mediated by binding of membrane-anchored transforming growth factor alpha to epidermal growth factor receptors promotes cell proliferation". Proceedings of the National Academy of Sciences of the United States of America. 87 (9): 3289–93. Bibcode:1990PNAS...87.3289A. doi:10.1073/pnas.87.9.3289. PMC 53885. PMID 2333283.
- Gilbert, Scott F. (2000). "Juxtacrine Signaling". In NCBI bookshelf (ed.). Developmental biology (6. ed.). Sunderland, Mass.: Sinauer Assoc. ISBN 0878932437.
- Gilbert, Scott F. (2000). "Juxtacrine Signaling". Developmental Biology. 6th Edition.
- Wells, Alan; Wiley, H. Steven (26 October 2018). "A systems perspective of heterocellular signaling". Essays in Biochemistry. 62 (4): 607–617. doi:10.1042/EBC20180015. ISSN 1744-1358. PMC 6309864. PMID 30139877.
- Nikolov, Dimitar B.; Xu, Kai; Himanen, Juha P. (October 2013). "Eph/ephrin recognition and the role of Eph/ephrin clusters in signaling initiation". Biochimica et Biophysica Acta (BBA) - Proteins and Proteomics. 1834 (10): 2160–2165. doi:10.1016/j.bbapap.2013.04.020. ISSN 0006-3002. PMC 3777820. PMID 23628727.
- Crawford, KM; Zambryski, PC (October 1999). "Plasmodesmata signaling: many roles, sophisticated statutes" (PDF). Current Opinion in Plant Biology. 2 (5): 382–7. doi:10.1016/s1369-5266(99)00009-6. PMID 10508755.
- Bruce Alberts; et al. (2002). "General Principles of Cell Communication". In NCBI bookshelf (ed.). Molecular biology of the cell (4th ed.). New York: Garland Science. ISBN 0815332181.
- Ding, Biao; Itaya, Asuka; Woo, Young-Min (1 January 1999), Jeon, Kwang W. (ed.), "Plasmodesmata and Cell-to-Cell Communication in Plants", International Review of Cytology, Academic Press, vol. 190, pp. 251–316, retrieved 27 February 2022
- Giancotti, FG; Ruoslahti, E (13 August 1999). "Integrin signaling". Science. 285 (5430): 1028–32. doi:10.1126/science.285.5430.1028. PMID 10446041.
External links
- "Autocrine versus juxtacrine signaling modes" - illustration at sysbio.org