Imd pathway
The Imd pathway is a broadly-conserved NF-κB immune signalling pathway of insects and some arthropods[1] that regulates a potent antibacterial defence response. The pathway is named after the discovery of a mutation causing severe immune deficiency (the gene was named "Imd" for "immune deficiency"). The Imd pathway was first discovered in 1995 using Drosophila fruit flies by Bruno Lemaitre and colleagues, who also later discovered that the Drosophila Toll gene regulated defence against Gram-positive bacteria and fungi.[2][3] Together the Toll and Imd pathways have formed a paradigm of insect immune signalling; as of September 2, 2019, these two landmark discovery papers have been cited collectively over 5000 times since publication on Google Scholar.[4][5]
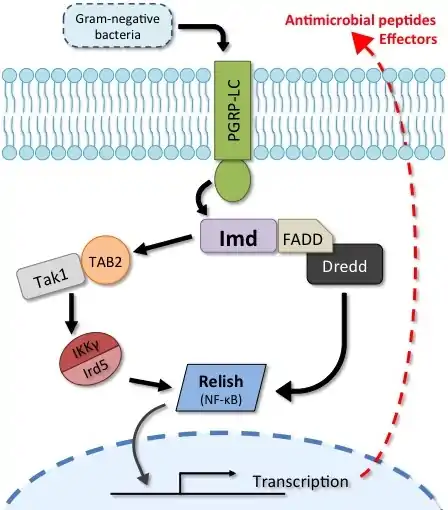
The Imd pathway responds to signals produced by Gram-negative bacteria. Peptidoglycan recognition proteins (PGRPs) sense DAP-type peptidoglycan, which activates the Imd signalling cascade. This culminates in the translocation of the NF-κB transcription factor Relish, leading to production of antimicrobial peptides and other effectors.[6] Insects lacking Imd signalling either naturally or by genetic manipulation are extremely susceptible to infection by a wide variety of pathogens and especially bacteria.
Similarity to human pathways
The Imd pathway bears a number of similarities to mammalian TNFR signalling, though many of the intracellular regulatory proteins of Imd signalling also bear homology to different signalling cascades of human Toll-like receptors.[6]
Similarity to TNFR signalling
The following genes are analogous or homologous between Drosophila melanogaster (in bold) and human TNFR1 signalling:[7][8]
- Imd: human orthologue = RIP1
- Tak1: human orthologue = Tak1
- TAB2: human orthologue = TAB2
- Dredd: human orthologue = caspase-8
- FADD: human orthologue = FADD
- Key/Ikkγ: human orthologue = NEMO[8]
- Ird5: human orthologue = IKK2
- Relish: human orthologues = p65/p50 and IκB
- Iap2: human orthologue = cIAP2
- UEV1a: human orthologue = UEV1a
- bend: human orthologue = UBC13
In Drosophila
While the exact epistasis of Imd pathway signalling components is continually scrutinized, the mechanistic order of many key components of the pathway is well-established. The following sections discuss Imd signalling as it is found in Drosophila melanogaster, where it is exceptionally well-characterized.[6] Imd signalling is activated by a series of steps from recognition of a bacterial substance (e.g. peptidoglycan) to the transmission of that signal leading to activation of the NF-κB transcription factor Relish.[7] Activated Relish then forms dimers that move into the nucleus and bind to DNA leading to the transcription of antimicrobial peptides and other effectors.
Peptidoglycan recognition proteins (PGRPs)
The sensing of bacterial signals is performed by peptidoglycan recognition protein LC (PGRP-LC), a transmembrane protein with an intracellular domain. Binding of bacterial peptidoglycan leads to dimerization of PGRP-LC which generates the conformation needed to bind and activate the Imd protein. However alternate isoforms of PGRP-LC can also be expressed with different functions: PGRP-LCx recognizes polymeric peptidoglycan, while PGRP-LCa does not bind peptidoglycan directly but acts alongside PGRP-LCx to bind monomeric peptidoglycan fragments (called tracheal cytotoxin or "TCT"). Another PGRP (PGRP-LE) also acts intracellularly to bind TCT that has crossed the cell membrane or is derived from an intracellular infection. PGRP-LA promotes the activation of Imd signalling in epithelial cells, but the mechanism is still unknown.[6][7]
Other PGRPs can inhibit the activation of Imd signalling by binding bacterial signals or inhibiting host signalling proteins: PGRP-LF is a transmembrane PGRP that lacks an intracellular domain and does not bind peptidoglycan. Instead PGRP-LF forms dimers with PGRP-LC preventing PGRP-LC dimerization and consequently activation of Imd signalling. A number of secreted PGRPs have amidase activity that downregulate the Imd pathway by digesting peptidoglycan into short, non-immunogenic fragments. These include PGRP-LB, PGRP-SC1A, PGRP-SC1B, and PGRP-SC2. Additionally, PGRP-LB is the major regulator in the gut.[9]
Intracellular signalling components

The principle intracellular signalling protein is Imd, a death domain-containing protein that binds with FADD and Dredd to form a complex. Dredd is activated following ubiquitination by the Iap2 complex (involving Iap2, UEV1a, bend, and eff), which allows Dredd to cleave the 30 residue N-terminus of Imd, allowing it to also be ubiquitinated by Iap2.[7] Following this, the Tak1/TAB2 complex binds to the activated form of Imd and subsequently activates the IKKγ/Ird5 complex through phosphorylation. This IKKγ complex activates Relish by phosphorylation, leading to cleavage of Relish and thereby producing both N-terminal and C-terminal Relish fragments. The N-terminal Relish fragments dimerize leading to their translocation into the nucleus where these dimers bind to Relish-family NF-κB binding sites. Binding of Relish promotes the transcription of effectors such as antimicrobial peptides.[6][7]
While Relish is integral for transcription of Imd pathway effectors, there is additional cooperation with other pathways such as Toll and JNK. The TAK1/TAB2 complex is key to propagating intracellular signalling of not only the Imd pathway, but also the JNK pathway. As a result, mutants for JNK signalling have severely reduced expression of Imd pathway antimicrobial peptides.[10]
The antimicrobial response
Imd signalling regulates a number of effector peptides and proteins that are produced en masse following immune challenge.[11] This includes many of the major antimicrobial peptide genes of Drosophila, particularly: Diptericin, Attacin, Drosocin, Cecropin, and Defensin.[12] The Imd pathway regulates hundreds of genes after infection, however the antimicrobial peptides play one of the most essential roles of Imd signalling in defence. Flies lacking multiple antimicrobial peptide genes succumb to infections by a broad suite of Gram-negative bacteria.[13][14] Classical thinking suggested that antimicrobial peptides worked as a generalist cocktail in defence, where each peptide provided a small and somewhat redundant contribution.[15][16] However Hanson and colleagues found that single antimicrobial peptide genes displayed an unexpectedly high degree of specificity for defence against specific microbes.[13] The fly Diptericin A gene is essential for defence against the bacterium Providencia rettgeri (also suggested by an earlier evolutionary study[17]). A second specificity is encoded by Diptericin B, which defends flies against Acetobacter bacteria of the fly microbiome.[18] A third specificity is encoded by the gene Drosocin. Flies lacking Drosocin are highly susceptible to Enterobacter cloacae infection.[13][14][19] The Drosocin gene itself encodes two peptides (named Drosocin and Buletin), wherein it is specifically the Drosocin peptide that is responsible for defence against E. cloacae, while the Buletin peptide instead mediates a specific defence against another bacterium, Providencia burhodogranariea.[19] These works accompany others on antimicrobial peptides and effectors regulated by the Drosophila Toll pathway, which also display a specific importance in defence against certain fungi or bacteria.[20][21][22]
This work on Drosophila immune antimicrobial peptides and effectors has greatly revised the former view that such peptides are generalist molecules. The modern interpretation is now that specific molecules might provide a somewhat redundant layer of defence, but also single peptides can have critical importance, individually, against relevant microbes.[23][24][25][26]
Conservation in insects
-PLoS.jpg.webp)
The Imd pathway appears to have evolved in the last common ancestor of centipedes and insects.[1] However certain lineages of insects have since lost core components of Imd signalling. The first-discovered and most famous example is the pea aphid Acyrthosiphon pisum. It is thought that plant-feeding aphids have lost Imd signalling as they bear a number of bacterial endosymbionts, including both nutritional symbionts that would be disrupted by aberrant expression of antimicrobial peptides, and defensive symbionts that cover for some of the immune deficiency caused by loss of Imd signalling.[27] It has also been suggested that antimicrobial peptides, the downstream components of Imd signalling, may be detrimental to fitness and lost by insects with exclusively plant-feeding ecologies.[28]
Crosstalk between the Imd and Toll signalling pathways
While the Toll and Imd signalling pathways of Drosophila are commonly depicted as independent for explanatory purposes, the underlying complexity of Imd signalling involves a number of likely mechanisms wherein Imd signalling interacts with other signalling pathways including Toll and JNK.[6] While the paradigm of Toll and Imd as largely independent provides a useful context for the study of immune signalling, the universality of this paradigm as it applies to other insects has been questioned. In Plautia stali stinkbugs, suppression of either Toll or Imd genes simultaneously leads to reduced activity of classic Toll and Imd effectors from both pathways.[29]
Insects and arthropods lacking Imd signalling
- The pea aphid Acyrthosiphon pisum[27]
- The bed bug Cimex lectularius[30]
- The mite Tetranychus urticae[31]
References
- Palmer WJ, Jiggins FM (August 2015). "Comparative Genomics Reveals the Origins and Diversity of Arthropod Immune Systems". Molecular Biology and Evolution. 32 (8): 2111–2129. doi:10.1093/molbev/msv093. PMC 4833078. PMID 25908671.
- Lemaitre B, Kromer-Metzger E, Michaut L, Nicolas E, Meister M, Georgel P, et al. (October 1995). "A recessive mutation, immune deficiency (imd), defines two distinct control pathways in the Drosophila host defense". Proceedings of the National Academy of Sciences of the United States of America. 92 (21): 9465–9469. Bibcode:1995PNAS...92.9465L. doi:10.1073/pnas.92.21.9465. PMC 40822. PMID 7568155.
- Lemaitre B, Nicolas E, Michaut L, Reichhart JM, Hoffmann JA (September 1996). "The dorsoventral regulatory gene cassette spätzle/Toll/cactus controls the potent antifungal response in Drosophila adults". Cell. 86 (6): 973–983. doi:10.1016/s0092-8674(00)80172-5. PMID 8808632. S2CID 10736743.
- "A recessive mutation, immune deficiency (imd), defines two distinct control pathways in the Drosophila host defense". Google Scholar. Retrieved 2 September 2019.
- "The dorsoventral regulatory gene cassette spätzle/Toll/cactus controls the potent antifungal response in Drosophila adults". Google Scholar. Retrieved 2 September 2019.
- Lemaitre B, Hoffmann J (2007). "The host defense of Drosophila melanogaster". Annual Review of Immunology. 25: 697–743. doi:10.1146/annurev.immunol.25.022106.141615. PMID 17201680.
- Myllymäki H, Valanne S, Rämet M (April 2014). "The Drosophila imd signaling pathway". Journal of Immunology. 192 (8): 3455–3462. doi:10.4049/jimmunol.1303309. PMID 24706930.
- "UniProtKB - Q9GYV5 (NEMO_DROME)". Uniprot.org.
Interpro family: IPR034735 NEMO_ZF
- Zaidman-Rémy A, Hervé M, Poidevin M, Pili-Floury S, Kim MS, Blanot D, et al. (April 2006). "The Drosophila amidase PGRP-LB modulates the immune response to bacterial infection". Immunity. 24 (4): 463–473. doi:10.1016/j.immuni.2006.02.012. PMID 16618604.
- Delaney JR, Stöven S, Uvell H, Anderson KV, Engström Y, Mlodzik M (July 2006). "Cooperative control of Drosophila immune responses by the JNK and NF-kappaB signaling pathways". The EMBO Journal. 25 (13): 3068–3077. doi:10.1038/sj.emboj.7601182. PMC 1500970. PMID 16763552.
- De Gregorio E, Spellman PT, Tzou P, Rubin GM, Lemaitre B (June 2002). "The Toll and Imd pathways are the major regulators of the immune response in Drosophila". The EMBO Journal. 21 (11): 2568–2579. doi:10.1093/emboj/21.11.2568. PMC 126042. PMID 12032070.
- Imler JL, Bulet P (2005). Antimicrobial peptides in Drosophila: structures, activities and gene regulation. Chemical Immunology and Allergy. Vol. 86. pp. 1–21. doi:10.1159/000086648. ISBN 3-8055-7862-8. PMID 15976485.
- Hanson MA, Dostálová A, Ceroni C, Poidevin M, Kondo S, Lemaitre B (February 2019). "Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach". eLife. 8: e44341. doi:10.7554/eLife.44341. PMC 6398976. PMID 30803481.
- Carboni AL, Hanson MA, Lindsay SA, Wasserman SA, Lemaitre B (January 2022). "Cecropins contribute to Drosophila host defense against a subset of fungal and Gram-negative bacterial infection". Genetics. 220 (1): iyab188. doi:10.1093/genetics/iyab188. PMC 8733632. PMID 34791204.
- Lazzaro BP (June 2008). "Natural selection on the Drosophila antimicrobial immune system". Current Opinion in Microbiology. 11 (3): 284–289. doi:10.1016/j.mib.2008.05.001. PMC 2527063. PMID 18555739.
- Lemaitre B, Hoffmann J (2007). "The host defense of Drosophila melanogaster". Annual Review of Immunology. 25: 697–743. doi:10.1146/annurev.immunol.25.022106.141615. PMID 17201680.
- Unckless RL, Howick VM, Lazzaro BP (January 2016). "Convergent Balancing Selection on an Antimicrobial Peptide in Drosophila". Current Biology. 26 (2): 257–262. doi:10.1016/j.cub.2015.11.063. PMC 4729654. PMID 26776733.
- Hanson, M. A.; Grollmus, L.; Lemaitre, B. (2023-07-21). "Ecology-relevant bacteria drive the evolution of host antimicrobial peptides in Drosophila". Science. 381 (6655). doi:10.1126/science.adg5725. ISSN 0036-8075.
- Hanson MA, Kondo S, Lemaitre B (June 2022). "Drosophila immunity: the Drosocin gene encodes two host defence peptides with pathogen-specific roles". Proceedings. Biological Sciences. 289 (1977): 20220773. doi:10.1098/rspb.2022.0773. PMC 9233930. PMID 35730150.
- Clemmons AW, Lindsay SA, Wasserman SA (April 2015). Silverman N (ed.). "An effector Peptide family required for Drosophila toll-mediated immunity". PLOS Pathogens. 11 (4): e1004876. doi:10.1371/journal.ppat.1004876. PMC 4411088. PMID 25915418.
- Cohen LB, Lindsay SA, Xu Y, Lin SJ, Wasserman SA (2020). "The Daisho Peptides Mediate Drosophila Defense Against a Subset of Filamentous Fungi". Frontiers in Immunology. 11: 9. doi:10.3389/fimmu.2020.00009. PMC 6989431. PMID 32038657.
- Hanson MA, Cohen LB, Marra A, Iatsenko I, Wasserman SA, Lemaitre B (August 2021). Lin X (ed.). "The Drosophila Baramicin polypeptide gene protects against fungal infection". PLOS Pathogens. 17 (8): e1009846. doi:10.1371/journal.ppat.1009846. PMC 8423362. PMID 34432851.
- Lin SJ, Cohen LB, Wasserman SA (May 2020). Silverman N (ed.). "Effector specificity and function in Drosophila innate immunity: Getting AMPed and dropping Boms". PLOS Pathogens. 16 (5): e1008480. doi:10.1371/journal.ppat.1008480. PMC 7255597. PMID 32463841.
- Hanson MA, Lemaitre B (February 2020). "New insights on Drosophila antimicrobial peptide function in host defense and beyond". Current Opinion in Immunology. 62: 22–30. doi:10.1016/j.coi.2019.11.008. PMID 31835066. S2CID 209357523.
- Lazzaro BP, Zasloff M, Rolff J (May 2020). "Antimicrobial peptides: Application informed by evolution". Science. 368 (6490): eaau5480. doi:10.1126/science.aau5480. PMC 8097767. PMID 32355003.
- Bosch TC, Zasloff M (October 2021). "Antimicrobial Peptides-or How Our Ancestors Learned to Control the Microbiome". mBio. 12 (5): e0184721. doi:10.1128/mBio.01847-21. PMC 8546549. PMID 34579574.
- Gerardo NM, Altincicek B, Anselme C, Atamian H, Barribeau SM, de Vos M, et al. (2010). "Immunity and other defenses in pea aphids, Acyrthosiphon pisum". Genome Biology. 11 (2): R21. doi:10.1186/gb-2010-11-2-r21. PMC 2872881. PMID 20178569.
- Hanson MA, Lemaitre B, Unckless RL (2019). "Dynamic Evolution of Antimicrobial Peptides Underscores Trade-Offs Between Immunity and Ecological Fitness". Frontiers in Immunology. 10: 2620. doi:10.3389/fimmu.2019.02620. PMC 6857651. PMID 31781114.
- Nishide Y, Kageyama D, Yokoi K, Jouraku A, Tanaka H, Futahashi R, Fukatsu T (February 2019). "Functional crosstalk across IMD and Toll pathways: insight into the evolution of incomplete immune cascades". Proceedings. Biological Sciences. 286 (1897): 20182207. doi:10.1098/rspb.2018.2207. PMC 6408883. PMID 30963836.
- Benoit JB, Adelman ZN, Reinhardt K, Dolan A, Poelchau M, Jennings EC, et al. (February 2016). "Unique features of a global human ectoparasite identified through sequencing of the bed bug genome". Nature Communications. 7: 10165. Bibcode:2016NatCo...710165B. doi:10.1038/ncomms10165. PMC 4740739. PMID 26836814.
- Santos-Matos G, Wybouw N, Martins NE, Zélé F, Riga M, Leitão AB, et al. (June 2017). "Tetranychus urticae mites do not mount an induced immune response against bacteria". Proceedings. Biological Sciences. 284 (1856): 20170401. doi:10.1098/rspb.2017.0401. PMC 5474072. PMID 28592670.