Integrin-linked kinase
Integrin-linked kinase is an enzyme that in humans is encoded by the ILK gene involved with integrin-mediated signal transduction. Mutations in ILK are associated with cardiomyopathies.[5][6] It is a 59kDa protein originally identified in a yeast-two hybrid screen with integrin β1 as the bait protein.[7] Since its discovery, ILK has been associated with multiple cellular functions including cell migration, proliferation, and adhesion.
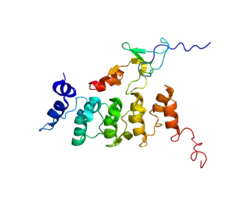




Integrin-linked kinases (ILKs) are a subfamily of Raf-like kinases (RAF). The structure of ILK consists of three features: 5 ankyrin repeats in the N-terminus, Phosphoinositide binding motif and extreme N-terminus of kinase catalytic domain.[8] Integrins lack enzymatic activity and depend on adapters to signal proteins.[8] ILK is linked to beta-1 and beta-3 integrin cytoplasmic domains and is one of the best described integrins.[9] Although first described as a serine/threonine kinase by Hannigan,[7] important motifs of ILK kinases are still uncharacterized.[9] ILK is thought to have a role in development regulation and tissue homeostasis, however it was found that in flies, worms and mice ILK activity isn't required to regulate these processes.[9]
Animal ILKs have been linked to the pinch- parvin complex which control muscle development.[9] Mice lacking ILK were embryonic lethal due to lack of organized muscle cell development.[9] In mammals ILK lacks catalytic activity but supports scaffolding protein functions for focal adhesions.[9] In plants, ILKs signal complexes to focal adhesion sites.[10] ILKs of plants contain multiple ILK genes. Unlike animals that contain few ILK genes[10] ILKs have been found to possess oncogenic properties. ILKs control the activity of serine/threonine phosphatases.[9]
Principle Features
Transduction of extracellular matrix signals through integrins influences intracellular and extracellular functions, and appears to require interaction of integrin cytoplasmic domains with cellular proteins. Integrin-linked kinase (ILK), interacts with the cytoplasmic domain of beta-1 integrin. Multiple alternatively spliced transcript variants encoding the same protein have been found for this gene.[11] Recent results showed that the C-terminal kinase domain is actually a pseudo-kinase with adaptor function.[12][13][14]
In 2008, ILK was found to localize to the centrosome and regulate mitotic spindle organization.[15]
Integrin-linked kinase has been shown to interact with:
Function of Plant ILK1
ILKs function by interacting with the many transmembrane receptors to regulate different signaling cascades.[7] ILK1 has been found in the root system of most plants where they are co-localized on the plasma membrane and endoplasmic reticulum where they transport ions across the plasma membrane[10] ILK1 is responsible for the control of osmotic and salt stress, control of the uptake of nutrients based on availability and pathogen detection.[23]
Osmotic and salt stress
ILK1 is linked to hyperosmotic stress sensitivity.[23] ILK1 reduced salt stress in seedlings placed in solution with increased concentrations of salt.[10] ILK1 concentrations remain fairly constant throughout development regardless of a high salt exposure.[23] Previously, it was believed that K+ accumulation was reduced in increased salt concentration.[24] K+ homeostasis is not affected in high salt concentrations. During periods of high salt stress, K+ concentrations in the presence of ILK1 was maintained at the existing level. Potassium transport is required for flg22 root growth inhibition and potassium transport was affected by flg22.[23]
Potassium levels modulate the activation of flg22, a flagellin peptide composed of 22 amino acids that triggers pathogen-associated molecular patterns (PAMPs). PAMPs functions by activating regulators of bacterial pathogen alert system.[23][25] Ion concentration levels of Mn2+, Mg2+, S and Ca2+ were also affected after PAMP regulators were mobilized.[23]
Nutrient uptake
Potassium (K+) is responsible for osmoregulation, membrane potential maintenance and turgor pressure of plant cells which in turn mediates stomata movement and growth of tubules within the plant.[26] Photosynthesis and other metabolic pathways are controlled by potassium.[26] When sufficient K+ uptake is not met, PAMPs are activated. Calmodulins, specifically CML9, have appeared as important genes to interact with ILK1 and regulate potassium levels within the cell. While CLM9 primarily regulates Ca2+ it is linked to a yet identified K+/Ca2+ influx channel.[10] While interactions are known to occur between CML9 and ILK1, ILK1 Is not a direct phosphorylation target of CML9. With the addition of CML9, autophosphorylation of ILK1 is diminished, the present irrespective of calcium available for uptake.

ILK1 is also affected by presence or absence of manganese (Mn2+). Autophosphorylation and substrate phosphorylation occurred when exposed to both Mn2+ and Mg2+. Mn2+ and was dose dependent where Mg2+ was not. Specific ILK autophosphorylation sites were found in the presence of Mn2+ but not in the presence of Mg2+ which supports the ILK1 dependent phosphorylation suggested above.[10] Mass spectrometry revealed no other kinases were present to trigger this response.
Pathogen detection
ILK1 has been found to promote resistance in bacterial pathogens.[10] ILK1 is required for flg22 sensitivity in seedlings. A catalytically inactive version of ILK1 was compared with catalytically active versions of ILK1 to see the level of resistance when challenged with bacterial pathogens. Plants inoculated with inactive ILK1 were more susceptible to bacterial infection than active ILK1 suggesting that ILK1 is needed for bacterial pathogen detection. While ILK1 is involved in bacterial pathogen detection it is not used for effect induced defenses.[23]
ILK1 increases PAMP response and basal immunity through phosphorylation of MPK3 and MPK6 and operates independently in reactive oxygen species (ROS) production. High Affinity Potassium uptake mediators such as HAK5 have also been found to be integral in the signaling of flg22.[23] HAK5 function when potassium levels are low.[23] Flg22 has been shown to depolarize the cell's plasma membrane with HAK5 and ILK1 working together to mediate ion homeostasis to assist with both short and long term actions such as growth and suppression thereof.[23]
References
- GRCh38: Ensembl release 89: ENSG00000166333 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000030890 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- Brodehl, Andreas; Rezazadeh, Saman; Williams, Tatjana; Munsie, Nicole M.; Liedtke, Daniel; Oh, Tracey; Ferrier, Raechel; Shen, Yaoqing; Jones, Steven J. M. (2019-02-15). "Mutations in ILK, encoding integrin-linked kinase, are associated with arrhythmogenic cardiomyopathy". Translational Research. 208: 15–29. doi:10.1016/j.trsl.2019.02.004. ISSN 1878-1810. PMC 7412573. PMID 30802431.
- Knöll, Ralph; Postel, Ruben; Wang, Jianming; Krätzner, Ralph; Hennecke, Gerrit; Vacaru, Andrei M.; Vakeel, Padmanabhan; Schubert, Cornelia; Murthy, Kenton (2007-07-31). "Laminin-alpha4 and integrin-linked kinase mutations cause human cardiomyopathy via simultaneous defects in cardiomyocytes and endothelial cells". Circulation. 116 (5): 515–525. doi:10.1161/CIRCULATIONAHA.107.689984. ISSN 1524-4539. PMID 17646580.
- Hannigan GE, Leung-Hagesteijn C, Fitz-Gibbon L, Coppolino MG, Radeva G, Filmus J, Bell JC, Dedhar S (1996). "Regulation of cell adhesion and anchorage-dependent growth by a new beta 1-integrin-linked protein kinase". Nature. 379 (6560): 91–6. Bibcode:1996Natur.379...91H. doi:10.1038/379091a0. PMID 8538749. S2CID 4325637.
- Dedhar S, Williams B, Hannigan G (1999). "Integrin-linked kinase (ILK): a regulator of integrin and growth-factor signalling". Trends in Cell Biology. 9 (8): 319–23. doi:10.1016/s0962-8924(99)01612-8. PMID 10407411.
- Widmaier M, Rognoni E, Radovanac K, Azimifar SB, Fässler R (2012). "Integrin-linked kinase at a glance". Journal of Cell Science. 125 (Pt 8): 1839–43. doi:10.1242/jcs.093864. PMID 22637643.
- Popescu SC, Brauer EK, Dimlioglu G, Popescu GV (2017). "Insights into the Structure, Function, and Ion-Mediated Signaling Pathways Transduced by Plant Integrin-Linked Kinases". Frontiers in Plant Science. 8: 376. doi:10.3389/fpls.2017.00376. PMC 5376563. PMID 28421082.
- "Entrez Gene: ILK integrin-linked kinase".
- Lange A, Wickström SA, Jakobson M, Zent R, Sainio K, Fässler R (October 2009). "Integrin-linked kinase is an adaptor with essential functions during mouse development". Nature. 461 (7266): 1002–6. Bibcode:2009Natur.461.1002L. doi:10.1038/nature08468. PMID 19829382. S2CID 4399882.
- Fukuda K, Gupta S, Chen K, Wu C, Qin J (December 2009). "The pseudoactive site of ILK is essential for its binding to alpha-Parvin and localization to focal adhesions". Molecular Cell. 36 (5): 819–30. doi:10.1016/j.molcel.2009.11.028. PMC 2796127. PMID 20005845.
- Qin J, Wu C (October 2012). "ILK: a pseudokinase in the center stage of cell-matrix adhesion and signaling". Current Opinion in Cell Biology. 24 (5): 607–13. doi:10.1016/j.ceb.2012.06.003. PMC 3467332. PMID 22763012.
- Fielding AB, Dobreva I, McDonald PC, Foster LJ, Dedhar S (February 2008). "Integrin-linked kinase localizes to the centrosome and regulates mitotic spindle organization". The Journal of Cell Biology. 180 (4): 681–9. doi:10.1083/jcb.200710074. PMC 2265580. PMID 18283114.
- Ewing RM, Chu P, Elisma F, Li H, Taylor P, Climie S, McBroom-Cerajewski L, Robinson MD, O'Connor L, Li M, Taylor R, Dharsee M, Ho Y, Heilbut A, Moore L, Zhang S, Ornatsky O, Bukhman YV, Ethier M, Sheng Y, Vasilescu J, Abu-Farha M, Lambert JP, Duewel HS, Stewart II, Kuehl B, Hogue K, Colwill K, Gladwish K, Muskat B, Kinach R, Adams SL, Moran MF, Morin GB, Topaloglou T, Figeys D (2007). "Large-scale mapping of human protein-protein interactions by mass spectrometry". Molecular Systems Biology. 3: 89. doi:10.1038/msb4100134. PMC 1847948. PMID 17353931.
- Barry FA, Gibbins JM (April 2002). "Protein kinase B is regulated in platelets by the collagen receptor glycoprotein VI". The Journal of Biological Chemistry. 277 (15): 12874–8. doi:10.1074/jbc.M200482200. PMID 11825911.
- Delcommenne M, Tan C, Gray V, Rue L, Woodgett J, Dedhar S (September 1998). "Phosphoinositide-3-OH kinase-dependent regulation of glycogen synthase kinase 3 and protein kinase B/AKT by the integrin-linked kinase". Proceedings of the National Academy of Sciences of the United States of America. 95 (19): 11211–6. Bibcode:1998PNAS...9511211D. doi:10.1073/pnas.95.19.11211. PMC 21621. PMID 9736715.
- Persad S, Attwell S, Gray V, Mawji N, Deng JT, Leung D, Yan J, Sanghera J, Walsh MP, Dedhar S (July 2001). "Regulation of protein kinase B/Akt-serine 473 phosphorylation by integrin-linked kinase: critical roles for kinase activity and amino acids arginine 211 and serine 343". The Journal of Biological Chemistry. 276 (29): 27462–9. doi:10.1074/jbc.M102940200. PMID 11313365.
- Leung-Hagesteijn C, Mahendra A, Naruszewicz I, Hannigan GE (May 2001). "Modulation of integrin signal transduction by ILKAP, a protein phosphatase 2C associating with the integrin-linked kinase, ILK1". The EMBO Journal. 20 (9): 2160–70. doi:10.1093/emboj/20.9.2160. PMC 125446. PMID 11331582.
- Tu Y, Li F, Goicoechea S, Wu C (March 1999). "The LIM-only protein PINCH directly interacts with integrin-linked kinase and is recruited to integrin-rich sites in spreading cells". Molecular and Cellular Biology. 19 (3): 2425–34. doi:10.1128/mcb.19.3.2425. PMC 84035. PMID 10022929.
- Zhang Y, Chen K, Guo L, Wu C (October 2002). "Characterization of PINCH-2, a new focal adhesion protein that regulates the PINCH-1-ILK interaction, cell spreading, and migration". The Journal of Biological Chemistry. 277 (41): 38328–38. doi:10.1074/jbc.M205576200. PMID 12167643.
- Brauer E (June 2016). "The Raf-like Kinsase ILK1 and the High Affinity K+ Transporter HAK5 Are Required for Innate Immunity and Abiotic Stress Response". Plant Physiology. 171 (2): 1470–1484. doi:10.1104/pp.16.00035. PMC 4902592. PMID 27208244 – via American Society of Plant Biologists.
- Alemán F, Nieves-Cordones M, Martínez V, Rubio F (2011). "Root K(+) acquisition in plants: the Arabidopsis thaliana model". Plant & Cell Physiology. 52 (9): 1603–12. doi:10.1093/pcp/pcr096. PMID 21771865.
- Chinchilla D, Bauer Z, Regenass M, Boller T, Felix G (2006). "The Arabidopsis receptor kinase FLS2 binds flg22 and determines the specificity of flagellin perception". The Plant Cell. 18 (2): 465–76. doi:10.1105/tpc.105.036574. PMC 1356552. PMID 16377758.
- Wang Y, Wu WH (October 2017). "Regulation of potassium transport and signaling in plants". Current Opinion in Plant Biology. 39: 123–128. doi:10.1016/j.pbi.2017.06.006. PMID 28710919.
Further reading
- Dedhar S (April 2000). "Cell-substrate interactions and signaling through ILK". Current Opinion in Cell Biology. 12 (2): 250–6. doi:10.1016/S0955-0674(99)00083-6. PMID 10712922.
- Persad S, Dedhar S (December 2003). "The role of integrin-linked kinase (ILK) in cancer progression". Cancer and Metastasis Reviews. 22 (4): 375–84. doi:10.1023/A:1023777013659. PMID 12884912. S2CID 24434819.
- Srivastava D, Yu S (September 2006). "Stretching to meet needs: integrin-linked kinase and the cardiac pump". Genes & Development. 20 (17): 2327–31. doi:10.1101/gad.1472506. PMID 16951248.
- Hannigan GE, Bayani J, Weksberg R, Beatty B, Pandita A, Dedhar S, Squire J (May 1997). "Mapping of the gene encoding the integrin-linked kinase, ILK, to human chromosome 11p15.5-p15.4". Genomics. 42 (1): 177–9. doi:10.1006/geno.1997.4719. PMID 9177792.
- Li F, Liu J, Mayne R, Wu C (October 1997). "Identification and characterization of a mouse protein kinase that is highly homologous to human integrin-linked kinase". Biochimica et Biophysica Acta. 1358 (3): 215–20. doi:10.1016/S0167-4889(97)00089-X. PMID 9366252.
- Chung DH, Lee JI, Kook MC, Kim JR, Kim SH, Choi EY, Park SH, Song HG (August 1998). "ILK (beta1-integrin-linked protein kinase): a novel immunohistochemical marker for Ewing's sarcoma and primitive neuroectodermal tumour". Virchows Archiv. 433 (2): 113–7. doi:10.1007/s004280050225. PMID 9737788. S2CID 24150231.
- Tu Y, Li F, Goicoechea S, Wu C (March 1999). "The LIM-only protein PINCH directly interacts with integrin-linked kinase and is recruited to integrin-rich sites in spreading cells". Molecular and Cellular Biology. 19 (3): 2425–34. doi:10.1128/mcb.19.3.2425. PMC 84035. PMID 10022929.
- Feng J, Ito M, Ichikawa K, Isaka N, Nishikawa M, Hartshorne DJ, Nakano T (December 1999). "Inhibitory phosphorylation site for Rho-associated kinase on smooth muscle myosin phosphatase". The Journal of Biological Chemistry. 274 (52): 37385–90. doi:10.1074/jbc.274.52.37385. PMID 10601309.
- Janji B, Melchior C, Vallar L, Kieffer N (June 2000). "Cloning of an isoform of integrin-linked kinase (ILK) that is upregulated in HT-144 melanoma cells following TGF-beta1 stimulation". Oncogene. 19 (27): 3069–77. doi:10.1038/sj.onc.1203640. PMID 10871859. S2CID 8798909.
- Velyvis A, Yang Y, Wu C, Qin J (February 2001). "Solution structure of the focal adhesion adaptor PINCH LIM1 domain and characterization of its interaction with the integrin-linked kinase ankyrin repeat domain". The Journal of Biological Chemistry. 276 (7): 4932–9. doi:10.1074/jbc.M007632200. PMID 11078733.
- Matsumoto M, Ogawa W, Hino Y, Furukawa K, Ono Y, Takahashi M, Ohba M, Kuroki T, Kasuga M (April 2001). "Inhibition of insulin-induced activation of Akt by a kinase-deficient mutant of the epsilon isozyme of protein kinase C". The Journal of Biological Chemistry. 276 (17): 14400–6. doi:10.1074/jbc.M011093200. PMID 11278835.
- Nikolopoulos SN, Turner CE (June 2001). "Integrin-linked kinase (ILK) binding to paxillin LD1 motif regulates ILK localization to focal adhesions". The Journal of Biological Chemistry. 276 (26): 23499–505. doi:10.1074/jbc.M102163200. PMID 11304546.
- Tu Y, Huang Y, Zhang Y, Hua Y, Wu C (April 2001). "A new focal adhesion protein that interacts with integrin-linked kinase and regulates cell adhesion and spreading". The Journal of Cell Biology. 153 (3): 585–98. doi:10.1083/jcb.153.3.585. PMC 2190577. PMID 11331308.
- Yamaji S, Suzuki A, Sugiyama Y, Koide Y, Yoshida M, Kanamori H, Mohri H, Ohno S, Ishigatsubo Y (June 2001). "A novel integrin-linked kinase-binding protein, affixin, is involved in the early stage of cell-substrate interaction". The Journal of Cell Biology. 153 (6): 1251–64. doi:10.1083/jcb.153.6.1251. PMC 2192033. PMID 11402068.
- Chen R, Kim O, Yang J, Sato K, Eisenmann KM, McCarthy J, Chen H, Qiu Y (August 2001). "Regulation of Akt/PKB activation by tyrosine phosphorylation". The Journal of Biological Chemistry. 276 (34): 31858–62. doi:10.1074/jbc.C100271200. PMID 11445557.