Danger model
The danger model of the immune system proposes that it differentiates between components that are capable of causing damage, rather that distinguishing between self and non-self.
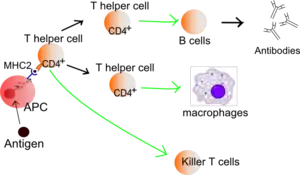
History of immunologic models
The first major immunologic model was the Self/Non-self Model proposed by Macfarlane Burnet and Frank Fenner in 1949 with later refinement by Burnet.[1][2] It theorizes that the immune system distinguishes between self, which is tolerated, and non-self, which is attacked and destroyed. According to this theory, the chief cell of the immune system is the B cell, activated by recognizing non-self structures. Later research showed that B cell activation is reliant on CD4+ T helper cells and a co-stimulatory signal from an antigen-presenting cell (APC). Because APCs are not antigen-specific, capable of processing self structures, Charles Janeway proposed the Infectious Non-self Model in 1989.[3] Janeway's theory involved APCs being activated by pattern recognition receptors (PRRs) that recognize evolutionarily conserved pathogen-associated molecular patterns (PAMPs) as infectious non-self, whereas PRRs are not activated by non-infectious self. However, neither of these models are sufficient to explain non-cytopathic viral infections, graft rejection, or anti-tumor immunity.[4]
Danger model
In 1994, Polly Matzinger formulated the danger model, theorizing that the immune system identifies threats to initiate an immune response based on the presence of pathogens and/or alarm signals from cells under stress.[5][6] When injured or stressed, tissues typically undergo non-silent types of cell death, such as necrosis or pyroptosis, releasing danger signals like DNA, RNA, heat shock proteins (Hsps), hyaluronic acid, serum amyloid A protein, ATP, uric acid, and cytokines like interferon-α, interleukin-1β, and CD40L for detection by dendritic cells.[4][6][7] In comparison, neoplastic tumors do not induce significant immune responses because controlled apoptosis degrades most danger signals, preventing the detection and destruction of malignant cells.[8]
Matzinger's work emphasizes that bodily tissues are the drivers of immunity, providing alarm signals on the location and extent of damage to minimize collateral damage.[9][10] The adaptive immune system relies on the innate immune system using its antigen-presenting cells to activate B and T lymphocytes for specific antibodies, exemplified by low dendritic cell counts resulting in common variable immunodeficiency (CVID).[11] For example, gut cells secrete transforming growth factor beta (TGF-β) during bacterial invasions to stimulate B cell production of Immunoglobulin A (IgA).[12] Similarly, 30-40% of the liver's T cells are Type I Natural Killer T (NTK) cells, providing Interleukin 4 (IL-4) for an organ-specific response of driving naïve CD4+ T cells to become Type 2 Helper T cells, as opposed to Type 1.[13][14]
Damage-associated molecular pattern (DAMP) model
Whereas the danger model proposes non-silent cell death releasing intracellular contents and/or expressing unique signalling proteins to stimulate an immune response, the damage-associated molecular pattern (DAMP) model theorizes that the immune system responds to exposed hydrophobic regions of biological molecules. In 2004, Seung-Yong Seong and Matzinger argued that as cellular damage causes denaturing and protein misfolding, exposed hydrophobic regions aggregate into clumps for improved binding to immune receptors.[15]
Pattern Recognition Receptors (PRRs)
Pattern Recognition Receptors (PRRs) are a family of surface receptors on antigen-presenting cells that includes toll-like receptors (TLRs), nucleotide oligomerization domain (NOD)-like receptors,[16] retinoic acid inducible gene-I (RIG-I)-like receptors[17] and C-type lectin-like receptors (CLRs).[18] They recognize alarmins, a category that includes both DAMPs and PAMPs, to process their antigenic regions for presentation to T helper cells.[6]
References
- Burnet FM, Fenner F (1949). The Production of Antibodies (2nd ed.). Melbourne: Macmillan.
- Burnet FM (1969). Cellular Immunology: Self and Notself. Cambridge: Cambridge University Press.
- Janeway CA (1989-01-01). "Approaching the asymptote? Evolution and revolution in immunology". Cold Spring Harbor Symposia on Quantitative Biology. 54 Pt 1 (1): 1–13. doi:10.1101/sqb.1989.054.01.003. PMID 2700931.
- Matzinger P (April 2002). "The danger model: a renewed sense of self". Science. 296 (5566): 301–305. Bibcode:2002Sci...296..301M. CiteSeerX 10.1.1.127.558. doi:10.1126/science.1071059. PMID 11951032. S2CID 13615808.
- Matzinger P (1994). "Tolerance, danger, and the extended family". Annual Review of Immunology. 12 (1): 991–1045. doi:10.1146/annurev.iy.12.040194.005015. PMID 8011301.
- Hallenbeck J, Del Zoppo G, Jacobs T, Hakim A, Goldman S, Utz U, Hasan A (December 2006). "Immunomodulation strategies for preventing vascular disease of the brain and heart: workshop summary". Stroke. 37 (12): 3035–3042. doi:10.1161/01.STR.0000248836.82538.ee. PMC 1853372. PMID 17082471.
- Jounai N, Kobiyama K, Takeshita F, Ishii KJ (2012). "Recognition of damage-associated molecular patterns related to nucleic acids during inflammation and vaccination". Frontiers in Cellular and Infection Microbiology. 2: 168. doi:10.3389/fcimb.2012.00168. PMC 3539075. PMID 23316484.
- Pradeu T, Cooper EL (2012-01-01). "The danger theory: 20 years later". Frontiers in Immunology. 3: 287. doi:10.3389/fimmu.2012.00287. PMC 3443751. PMID 23060876.
- Matzinger P (January 2007). "Friendly and dangerous signals: is the tissue in control?". Nature Immunology. 8 (1): 11–13. doi:10.1038/ni0107-11. PMID 17179963. S2CID 6448542.
- Matzinger P, Kamala T (March 2011). "Tissue-based class control: the other side of tolerance". Nature Reviews. Immunology. 11 (3): 221–230. doi:10.1038/nri2940. PMID 21350581. S2CID 10809131.
- Bayry J, Lacroix-Desmazes S, Kazatchkine MD, Galicier L, Lepelletier Y, Webster D, et al. (October 2004). "Common variable immunodeficiency is associated with defective functions of dendritic cells". Blood. 104 (8): 2441–2443. doi:10.1182/blood-2004-04-1325. PMID 15226176.
- Bauché D, Marie JC (April 2017). "Transforming growth factor β: a master regulator of the gut microbiota and immune cell interactions". Clinical & Translational Immunology. 6 (4): e136. doi:10.1038/cti.2017.9. PMC 5418590. PMID 28523126.
- Gao B, Jeong WI, Tian Z (February 2008). "Liver: An organ with predominant innate immunity". Hepatology. 47 (2): 729–736. doi:10.1002/hep.22034. PMID 18167066. S2CID 5441697.
- Yoshimoto T (2018). "The Hunt for the Source of Primary Interleukin-4: How We Discovered That Natural Killer T Cells and Basophils Determine T Helper Type 2 Cell Differentiation In Vivo". Frontiers in Immunology. 9: 716. doi:10.3389/fimmu.2018.00716. PMC 5924770. PMID 29740428.
- Seong S, Matzinger P (2004). "Hydrophobicity: an ancient damage-associated molecular pattern that initiates innate immune responses". Nature Reviews Immunology. 4 (6): 469–478. doi:10.1038/nri1372. PMID 15173835. S2CID 13336660.
- Tanti JF, Ceppo F, Jager J, Berthou F (2012). "Implication of inflammatory signaling pathways in obesity-induced insulin resistance". Front Endocrinol (Lausanne). 3: 181. doi:10.3389/fendo.2012.00181. PMC 3539134. PMID 23316186.
- Beckham SA, Brouwer J, Roth A, Wang D, et al. (2012). "Conformational rearrangements og RIG-I receptor on formation of a multiprotein: dsRNA assembly". Nucleic Acids Res. 41 (5): 3436–45. doi:10.1093/nar/gks1477. PMC 3597671. PMID 23325848.
- Kuroki K, Furukawa A, Maenaka K (2012). "Molecular recognition of paired receptors in the immune system". Front Microbiol. 3: 429. doi:10.3389/fmicb.2012.00429. PMC 3533184. PMID 23293633.