Casparian strip
The Casparian strip is a band-like thickening in the center of the root endodermis (radial and tangential walls of endodermal cells) of vascular plants (Pteridophytes [1] and Spermatophytes). The composition of the region is mainly suberin, lignin and some structural proteins, which are capable of reducing the diffusive apoplastic flow of water and solutes into the stele and its width varies between species.[2][3] The Casparian strip is impervious to water so can control the transportation of water and inorganic salts between the cortex and the vascular bundle, preventing water and inorganic salts from being transported to the stele through the apoplast, so that it must enter the cell membrane and move to the stele through the symplastic pathway, blocking the internal and external objects of the cell. The function and function of mass transportation are similar to that of animal tissues..[4][5] The development of the Casparian strip is regulated by transcription factors such as SHORT-ROOT (SHR), SCARECROW (SCR) and MYB36, as well as polypeptide hormone synthesised by midcolumn cells.[6][7]
.jpg.webp)
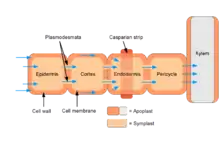
The chemistry of the Casparian strip has been described as composed of suberin. According to some studies,[8] the Casparian strip begins as a localized deposition of phenolic and unsaturated fatty substances in the middle lamella between the radial walls, as partly oxidized films. The primary wall becomes encrusted with and later thickened by deposits of similar substances on the inside of that wall. The encrustation of the cell wall by the material constituting the Casparian strip presumably plugs the pores that would have otherwise allowed the movement of water and nutrients via capillary action along that path.[9] The cytoplasm of the endodermal cell is firmly attached to the Casparian strip so that it does not readily separate from the strip when the cells are subjected to contraction of the protoplasts. At the root, the Casparian strip is embedded within the cell wall of endodermal cells in the non-growing region of the root behind the root tip.[10] Here, the Casparian strip serves as a boundary layer separating the apoplast of the cortex from the apoplast of the vascular tissue thereby blocking diffusion of material between the two.[11] This separation forces water and solutes to pass through the plasma membrane via a symplastic route in order to cross the endodermis layer.[10]
Casparian strips differentiate after an outward growth of the cortex is completed. At this level of the root development, the primary xylem of its vascular cylinder is only partly advanced. In gymnosperms and angiosperms displaying secondary growth, the roots commonly develop only endodermis with Casparian strips. In many of those, the endodermis is later discarded, together with the cortex, when the periderm develops from the pericycle. If the pericycle is superficial and the cortex is retained, either the endodermis is stretched or crushed or it keeps pace with the expansion of the vascular cylinder by radial anticlinal divisions, and the new walls develop Casparian strips in continuity with the old ones.[12]
In the absence of secondary growth (most monocotyledons and a few eudicots), the endodermis commonly undergoes wall modifications. There are two developmental stages beyond the development of the Casparian strip. In the second stage suberin (or endoderm[9]) coats the entire wall on the inside of the cell. As a result, the Casparian strip is separated from the cytoplasm and the connection between the two ceases to be evident. In the third stage, a thick cellulose layer is deposited over the suberin, sometimes mainly on the inner tangential walls. The thickened wall, as well as the original wall in which the Casparian strip is located, may become lignified, creating a secondary cell wall. The Casparian strip may be identifiable after the thickening of the endodermal wall has occurred. The thickened endodermal wall may have pits. The successive development of endodermal walls is clearly expressed in monocotyledons.[13][14]
Discovery
The discovery of the Casparian strip dates back to the mid-19th century, and advances in the understanding of the endodermis of plant roots.[15] In 1865, the German botanist Robert Caspary first described the endodermis of the root of plants, found that its cell wall was thickened, and named it Schuchtzscheide. Later scholars called the thickened part of it the Carls Belt, which was named after Casbury.[5][16] The term "Caspary'schen fleck" (German: Caspary'schen fleck) appeared in the 1870s literature,[17][18] and after the 20th century, it was often called the Casparian strip. In 1922, researchers first left the Casparian strip from the root of plants to study its composition. [19][20]
Composition
The chemical composition of the Casparian strip has been controversial for a long time. Casbury pointed out that this structure may be composed of lignin or suberin. Later scholars mostly thought it was suberin.[21] It was not until the 1990s that after analyzing the Casparian strip of several plants, it was found that lignin was the main component, but many textbooks have not been updated.[4] Although the cell wall of the endothelium is rich in woodbolic??(what does "woodbolic" even mean?suggest a better term), this is the result of the sublevel differentiation of the endothelium.[note 1] In the past, some scholars believe that the formation of the endodermis of Casparian strip is the beginning of sublevel differentiation, but there is no direct relationship between the two. The casparian strip has formed after the primary differentiation, and the secondary differentiation begins with the slash cut of the root, not where the Casparian strip is.[1]
Function
The casparian strip is fully filled with the gap between endothelial cells, including the middle lamella, making the cell walls of the two cells almost fused.[1] In the transportation of water and inorganic nutrients at the root of plants, the Casparian strip mainly affects the transportation of primary in vitro, that is, the transportation of water and inorganic salts through the interstitial cells of the epidermis and cortex cells. When water and inorganic salt come to the endothelial cells, they need to enter the cell through the cell membrane because the casparian strip is not water-permeable, and then transported by the protoplasmic inner path to reach the lignan cells of the stele, and then to other organs such as the stems and leaves.[16] When the growth environment is unfavourable, the casparian strip can act as a barrier between plant cells and the outside world, avoiding the entry of ions or outflow of their own ions in the environment.[7] In addition, the thickening of the carcass belt and the cortex also prevents toxic substances or pathogen invasion, as well as the function of preventing water dispersion.[22] Some studies have shown that plants may form thicker casparian strip in high-salt environments, and in areas closer to the tip of the roots, which may be an adaptation to the environment,[23][24] but compared with the endothelial sublevel differentiated wooden bolt walls, which are significantly thickened in high-salt adversity, the casparian strip changes is smaller.[25]
The casparian strip is mainly located in the endodermis of the root,[26] but some plants also have the casparian strip in the outer cortex on the outer side of the root cortex, stem or leaf.[27] For example, the conifers of Pinus bungeana and the stems of Pelargonium have the casparian strip, which may be related to preventing water dispersion or pathogenic invasion.[28][29]
Development
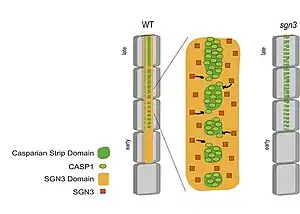
The development of Carters strap is started after the endogenic cells are fully delayed,[21][30] and there is currently two news signal transduction that promote endothelial cell formation of Casparian strip. The first is transcription factor Short-root (SHR) Activated additional two transcription factors Myb36 and Scarecrow (SCR), the former can stimulate Casparian Strip Proteins (Casp1-5), Peroxidase (PER64) and ESB1 (Enhanced) Suberin 1), etc., the latter, the latter affects the position of the carbide strap in the inner skin cell, which causes the position of the Casparian strip to be too close to the Stele;[6] the second is medium Casparian Strip Integrity Factor (CIF1-2) and the GSO1 (SGN3) and GSO2 receptor bonded to the endothelial cell radial wall and the GSO2 receptor in the lateral wall. CASP in the cells is concentrated to a cell membrane region corresponding to the position of the Cascade Band, forming a Casparian Strip Membrane Domain (CSD), and the CSD is incorporated in the region, The GS01 receptor is surrounded by the edge of each CSD region, promoting CSD fused into a continuous strip region, that is, the region where the Casparian strip is to be formed.[7][31]
Casparian strip protein is a membrane protein that interacts with each other and can bind to proteins needed to synthesize lignin such as PER64, ESB1 and respiratory oxidase homologer F (RBOHF) to activate the downstream reaction of Carzite band development.[1][5] In mutant plants lacking GSO1 receptors or at the same time lacking CIF1 and CIF2 polypeptides, CASP1 is abnormally distributed on the endothelial cell membrane, and the CSD cannot normally fuse into a continuous and complete band structure, thus eventually forming a broken and discontinuous Carlsian band.[7][31]
Environmental factors such as light, soil salinity and water deficit can affect the development of the Casparian strip.[28]
Photo
.jpg.webp) The vascular bundles of the root of the genus Acorus (monocotyledons) can be seen in the endothelium and the Casparian strip around the middle column. The Casparian strip is dyed red because it contains lignin.
The vascular bundles of the root of the genus Acorus (monocotyledons) can be seen in the endothelium and the Casparian strip around the middle column. The Casparian strip is dyed red because it contains lignin..jpg.webp) The vascular bundle of the root of the genus Ranunculus (dicotyledon) can see the endothelium and the Casparian strip around the middle column. The Casparian strip is dyed red because it contains lignin.
The vascular bundle of the root of the genus Ranunculus (dicotyledon) can see the endothelium and the Casparian strip around the middle column. The Casparian strip is dyed red because it contains lignin..jpg.webp) A Pinaceae (Gymnosperm) coniferous leaves. Peripheral endothelial endodermis and Casparian strip of visible vascular bundles
A Pinaceae (Gymnosperm) coniferous leaves. Peripheral endothelial endodermis and Casparian strip of visible vascular bundles
See also
Notes
- After the endothelial cell wall in the old root forms a wood embolism thickened, its function can be changed from transmission water to protecting plants, which can further limit the transmission of water and inorganic salts. Only channel cells (a few sublevel differentiated endothelial cells) retain transportation function. As the root grows, some plants lose channel cells in the root.[1]
References
- Geldner, N. (2013). "The Endodermis". Annual Review of Plant Biology. 64 (1): 531–558. doi:10.1146/annurev-arplant-050312-120050. ISSN 1543-5008. PMID 23451777.
- Chen, Tong; Cai, Xia; Wu, Xiaoqin; Karahara, Ichirou; Schreiber, Lucas; Lin, Jinxing (October 2011). "Casparian strip development and its potential function in salt tolerance". Plant Signaling & Behavior. 6 (10): 1499–1502. doi:10.4161/psb.6.10.17054. ISSN 1559-2316. PMC 3256377. PMID 21904117.
- Kirkham, M. B. (2005-01-01), Kirkham, M. B. (ed.), "14 - Root Anatomy and Poiseuille's Law for Water Flow in Roots", Principles of Soil and Plant Water Relations, Burlington: Academic Press, pp. 207–227, ISBN 978-0-12-409751-3, retrieved 2022-12-22
- Geldner, N. (2013). "Casparian strips" (PDF). Current Biology. Vol. 23, no. 23. pp. R1025, R1026.
- Roppolo, D.; De Rybel, B.; Tendon, V. D.; Pfister, A.; Alassimone, J.; Vermeer, J. E. M.; Yamazaki, M.; Stierhof, Y.-D.; Beeckman, T.; Geldner, N. (2011). "A novel protein family mediates Casparian strip formation in the endodermis". Nature. 473 (7347): 380–383. Bibcode:2011Natur.473..380R. doi:10.1038/nature10070. ISSN 0028-0836. PMID 21593871. S2CID 4366553.
- Li, P.; Yu, Q.; Gu, X.; Xu, C.; Qi, S.; Wang, H.; et al. (2018). "Construction of a Functional Casparian Strip in Non-endodermal Lineages Is Orchestrated by Two Parallel Signaling Systems in Arabidopsis thaliana". Current Biology. 28 (17): 2777–2786.e2. doi:10.1016/j.cub.2018.07.028. PMID 30057307.
- Nakayama, T.; Shinohara, H.; Tanaka, M.; Baba, K.; Ogawa-Ohnishi, M.; Matsubayashi, Y. (2017). "A peptide hormone required for Casparian strip diffusion barrier formation in Arabidopsis roots". Science. 355 (6322): 284–286. Bibcode:2017Sci...355..284N. doi:10.1126/science.aai9057. PMID 28104889. S2CID 206653442.
- Van Fleet, D. S. (1961). "Histochemistry and function of the endodermis". Botanical Review. 27 (2): 165–220. doi:10.1007/BF02860082. S2CID 45393531.
- Frey-Wyssling, A.; H. H. Bosshard (1959). Cytology of the Ray Cells in Sapwood and Heartwood. Cram.
- Taiz, L., Zeiger, Eduardo, Møller, Ian Max, & Murphy, Angus. (2015). Plant physiology and development (Sixth ed.).
- Grebe, M (2011). "Unveiling the Casparian strip". Nature. 473 (7347): 294–5. doi:10.1038/473294a. PMID 21593860. S2CID 873056.
- von Guttenberg, H. (1943). Die physiologischen Scheiden. Borntraeger.
- OGURA, Y. (1938). "Problems in morphology (13)". Botany and Zoology. 6: 139–148.
- Napp-Zinn, A. F. (1953). 100 Jahre Köln-düsseldorfer Rheindampfschiffahrt: Insbesondere Zerstörung und Wiederaufbau 1939-1953. Köln-Düsseldorfer Rheindampfshiffahrt.
- Seago, J. L., Jr. (2020). "Revisiting the occurrence and evidence of endodermis in angiosperm shoots". Flora. 273: 151709. doi:10.1016/j.flora.2020.151709.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Grebe, M. (2011). "Unveiling the Casparian strip". Nature. 473 (7347): 294–295. doi:10.1038/473294a. ISSN 0028-0836. PMID 21593860. S2CID 873056.
- Russow, E. (1872). "Rhizocarpeae. I. Axenorgane: A. Stamm: a. Marsilia (Drummondii, elata, salvatrix)". Vergleichende Untersuchungen betreffend die Histiologie (Histiographie und Histiogenie) der vegetativen und Sporen-bildenden Organe und die Entwickelung der Sporen der Leitbündel-Kryptogamen: mit Berücksichtigung der Histiologie der Phanerogamen, ausgehend von der Betrachtung der Marsiliaceen. Commissionnaires de l'Académie Impériale des sciences. pp. 1–12.
- Müller, C. (1884). "Morphologie der Gewebe". 12 (1): 234–342.
{{cite journal}}: Cite journal requires|journal=(help) - Priestley, J. H.; North, E. E. (1922). "Physiological Studies in Plant Anatomy III. The Structure of the Endodermis in Relation to its Function". The New Phytologist. 21 (3): 113–139. doi:10.1111/j.1469-8137.1922.tb07593.x. JSTOR 2428118.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Song, C.; Shen, W.; Du, L.; Wen, J.; Lin, J.; Li, R. (2019). "Development and chemical characterization of Casparian strips in the roots of Chinese fir (Cunninghamia lanceolata)". Trees. 33 (3): 827–836. doi:10.1007/s00468-019-01820-x. ISSN 0931-1890.
- Naseer, S.; Lee, Y.; Lapierre, C.; Franke, R.; Nawrath, C.; Geldner, N. (2012). "Casparian strip diffusion barrier in Arabidopsis is made of a lignin polymer without suberin". Proceedings of the National Academy of Sciences. 109 (25): 10101–6. Bibcode:2012PNAS..10910101N. doi:10.1073/pnas.1205726109. PMC 3382560. PMID 22665765.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Robbins, N. E.; Trontin, C.; Duan, L.; Dinneny, J. R. (2014). "Beyond the barrier: communication in the root through the endodermis". Plant Physiology. 166 (2): 551–9. doi:10.1104/pp.114.244871. PMC 4213087. PMID 25125504.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Karahara, I.; Ikeda, A.; Kondo, T.; Uetake, Y. (2004). "Development of the Casparian strip in primary roots of maize under salt stress". Planta. 219 (1): 41–7. doi:10.1007/s00425-004-1208-7. PMID 14986139. S2CID 792699.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Chen, T.; Cai, X.; Wu, X.; Karahara, I.; Schreiber, L.; Lin, J. (2011). "Casparian strip development and its potential function in salt tolerance". Plant Signaling & Behavior. 6 (10): 1499–502. doi:10.4161/psb.6.10.17054. PMC 3256377. PMID 21904117.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Palmgren, M. (2018). "Plant epithelia: What is the role of the mortar in the wall?". PLOS Biology. 16 (12): e3000073. doi:10.1371/journal.pbio.3000073. PMC 6296743. PMID 30517104.
- Evert, R. F. (2006-08-28). Esau's Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development. John Wiley & Sons. p. 6. ISBN 978-0-470-04737-8. Archived from the original on 2020-12-01. Retrieved 2021-04-17.
- Lersten, N. R. (1997). "Occurrence of endodermis with a casparian strip in stem and leaf". The Botanical Review. 63 (3): 265–272. doi:10.1007/BF02857952. ISSN 0006-8101. S2CID 45859773.
- Wu, X.; Lin, J.; Lin, Q.; Wang, J.; Schreiber, L. (2005). "Casparian Strips in Needles are More Solute Permeable than Endodermal Transport Barriers in Roots of Pinus bungeana". Plant and Cell Physiology. 46 (11): 1799–1808. doi:10.1093/pcp/pci194. ISSN 1471-9053. PMID 16170202.
- Meyer, C. J.; Peterson, C. A. (2011). "Casparian bands occur in the periderm of Pelargonium hortorum stem and root". Annals of Botany. 107 (4): 591–8. doi:10.1093/aob/mcq267. PMC 3064534. PMID 21239408.
{{cite journal}}: CS1 maint: multiple names: authors list (link) - Barberon, M. (2017). "The endodermis as a checkpoint for nutrients". New Phytologist. 213 (4): 1604–1610. doi:10.1111/nph.14140. PMID 27551946.
- Pfister, A.; Barberon, M.; Alassimone, J.; Kalmbach, L.; Lee, Y.; Vermeer, J. E. M.; et al. (2014). "A receptor-like kinase mutant with absent endodermal diffusion barrier displays selective nutrient homeostasis defects". eLife. 3: e03115. doi:10.7554/eLife.03115. PMC 4164916. PMID 25233277.
{{cite journal}}: CS1 maint: multiple names: authors list (link)
- Esau, Katherine (1965). Plant Anatomy. John Wiley & Sons. p. 767. ISBN 9780471244554.