ABHD18
ABHD18 is a protein that in Homo sapiens is encoded by the ABHD18 gene.[5]


| Chromosome 4 open reading frame 29 | |||||||
|---|---|---|---|---|---|---|---|
| Identifiers | |||||||
| Symbol | C4orf29 | ||||||
| NCBI gene | 80167 | ||||||
| HGNC | 26111 | ||||||
| RefSeq | NP_001034806.1 | ||||||
| UniProt | Q0P651 | ||||||
| Other data | |||||||
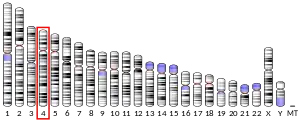
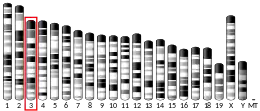
| Locus | Chr. 4 q28.2 | ||||||
| |||||||
Gene
ABHD18 is found on the positive strand of the human genome at 4q28.2. It is 74.4 kbp. The gene contains 17 exons.[6] The longest mRNA transcript is composed of 13 exons and is 2200 base pairs.[5]

Homology
Orthologs
Many orthologs to human ABHD18 have been discovered, with the most distant ortholog with high (over 90%) coverage is found in rice Oryza sativa.[8] The protein is not found in fungi. Bacteria of the order Myxobacteria and genus Chitinimonas contain orthologous regions to the C4orf29 protein. The few bacterial homologs indicate a horizontal gene transfer event. The domain of unknown function, DUF2048, is conserved throughout orthologs.
| Organism | NCBI Accession Number | Divergence from Humans (Million Years) | Protein Length (Amino Acids) | Sequence Identity to Human | Sequence Similarity to Human |
|---|---|---|---|---|---|
| Human | NP_001034806.1 | 0 | 414 | 100% | 100% |
| Dog | XP_005631879.1 | 94.2 | 484 | 87% | 89% |
| Canary | XP_009094557.1 | 296 | 414 | 75% | 87% |
| Painted turtle | XP_005282815.1 | 296 | 464 | 73% | 83% |
| Ant | XP_011141314.1 | 782 | 468 | 46% | 64% |
| Flat Worm | NP_492206.1 | 937 | 378 | 33% | 50% |
| Rice | NP_001046183.1 | 1369 | 366 | 31% | 47% |
| Cystobacter violaceus | WP_043404439.1 | 2535 | 321 | 31% | 52% |

Protein
ABHD18 codes a 414 amino acid sequence of 46.9 kDa in humans. The predicted isoelectric point is 9.37.[10] The domain of unknown function, DUF2048, is found from amino acid residues 25 to 414 in the precursor C4orf29 protein.[11] This domain is part of the alpha/beta hydrolase superfamily, which comprises enzymes that catalyze fat metabolism. Predicted post-translational modifications include glycosylation at residues Ser287 and Ser319 [12] and sumoylation[13] at the motifs Phe240 to Gly243, Ala377 to Asp340, and Phe408 to Gly411.
Expression
The protein product of ABHD18 in humans is predicted to be a secreted product. It is ubiquitously expressed at low to moderate levels.[14] In humans, the protein is found at high levels the digestive tract and parathyroid gland.[15] The homologous mouse protein 3110057O12Rik is expressed at high levels in the granule layer of the cerebellum.[16]
Clinical significance
ABHD18 contains highly variable numbers of Alu repeats.[17] A low number of Alu repeats in the human ABHD18 protein is associated with increase prevalence of hepatocellular carcinoma (HCC) in Asian populations. This information is used as a genetic marker to determine genetic risk of HCC.[18] Swine muscle transcriptome analysis indicates high expression of ABHD18 in swine with extreme low levels of fatty acid composition.[19]
References
- GRCh38: Ensembl release 89: ENSG00000164074 - Ensembl, May 2017
- GRCm38: Ensembl release 89: ENSMUSG00000037818 - Ensembl, May 2017
- "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- "C4orf29". NCBI Gene. NCBI. Retrieved 5 May 2015.
- "C4orf29". NCBI AceView. Retrieved 2015-05-05.
- "C4orf29". GeneCards Human Gene Database.
- "NP_001034806". NCBI Protein BLAST.
- Kelley, LA; Sternberg, MJE. "Protein structure prediction on the web: a case study using the Phyre server". Phyre2. Retrieved 16 Apr 2015.
- "C4orf29 (human)". Phosphosite.org. Retrieved 2015-05-05.
- "uncharacterized protein C4orf29 precursor [Homo sapiens]". NCBI Protein.
- "NetOGlyc4.0 C4orf29". CBS Prediction Servers.
- "C4orf29 Sumoylation". ABGENT.
- "Large-scale analysis of the human transcriptome GDS596". NCBI GeoProfile. Retrieved 2015-05-05.
- "C4orf29". The Human Protein Atlas. Retrieved 2015-05-05.
- "3110057O12Rik". Allen Brain Atlas. Retrieved 2015-05-05.
- Osenberg S, Paz Yaacov N, Safran M, Moshkovitz S, Shtrichman R, Sherf O, Jacob-Hirsch J, Keshet G, Amariglio N, Itskovitz-Eldor J, Rechavi G (June 2010). "Alu sequences in undifferentiated human embryonic stem cells display high levels of A-to-I RNA editing". PLOS ONE. 5 (6): e11173. Bibcode:2010PLoSO...511173O. doi:10.1371/journal.pone.0011173. PMC 2888580. PMID 20574523.
- Clifford RJ, Zhang J, Meerzaman DM, Lyu MS, Hu Y, Cultraro CM, Finney RP, Kelley JM, Efroni S, Greenblum SI, Nguyen CV, Rowe WL, Sharma S, Wu G, Yan C, Zhang H, Chung YH, Kim JA, Park NH, Song IH, Buetow KH (December 2010). "Genetic variations at loci involved in the immune response are risk factors for hepatocellular carcinoma". Hepatology. 52 (6): 2034–43. doi:10.1002/hep.23943. PMC 8259333. PMID 21105107. S2CID 5286278.
- Puig-Oliveras A, Ramayo-Caldas Y, Corominas J, Estellé J, Pérez-Montarelo D, Hudson NJ, Casellas J, Folch JM, Ballester M (June 2014). "Differences in muscle transcriptome among pigs phenotypically extreme for fatty acid composition". PLOS ONE. 9 (6): e99720. Bibcode:2014PLoSO...999720P. doi:10.1371/journal.pone.0099720. PMC 4057286. PMID 24926690.