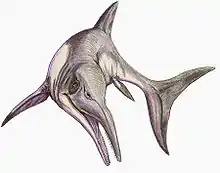
Besanosaurus
Besanosaurus (meaning 'reptile from Besano') is a genus of Middle Triassic ichthyosaur from Monte San Giorgio of Italy and Switzerland, containing the single species B. leptorhynchus. Besanosaurus was named by Cristiano Dal Sasso and Giovanni Pinna in 1996, based on the nearly complete flattened skeleton BES SC 999, the holotype specimen. This skeleton is preserved across multiple thin rock slabs spanning 3.5 by 4 metres (11 by 13 ft) when assembled and took thousands of hours to prepare. Additional specimens from Monte San Giorgio that have previously been considered separate genera, including a partial skull named Mikadocephalus and a well-preserved, largely complete skeleton, have been reinterpreted as additional specimens of Besanosaurus.
| Besanosaurus Temporal range: Middle Triassic, latest | |
|---|---|
 | |
| Skull and front part of the skeleton of the holotype | |
| Scientific classification | |
| Domain: | |
| Kingdom: | |
| Phylum: | |
| Class: | |
| Order: | |
| (unranked): | |
| Genus: | †Besanosaurus Dal Sasso and Pinna, 1996 |
| Species | |
| |
| Synonyms | |
| |
As an ichthyosaur, Besanosaurus had flippers for limbs and a fin on the tail. Besanosaurus is a large ichthyosaur, with the largest known specimen estimated to measure about 8 metres (26 ft) long. It has a long, slender body with a small head and long tail. The snout of Besanosaurus is long and thin, and contains numerous small pointed teeth. In the upper jaw, the teeth are mostly set into sockets but the rearmost teeth are implanted in a groove. The lower jaw bears enlarged coronoid processes for the anchorage of jaw muscles. There are 60 vertebrae in front of the hips, two in the hip region, and 139 in the tail. The scapulae are sickle-shaped, and the forelimbs are longer than the hindlimbs. The humeri are round and squat, and the finger bones are elliptical.
While it is understood to be a shastasaurid-type ichthyosaur, how exactly Besanosaurus and other members of this group are related to each other is unclear. The skull bones of Besanosaurus indicate that it would have possessed strong jaw muscles, but its delicate snout suggests it would have fed on small fish and coleoid cephalopods, which it could have caught with rapid, snapping bites. Ichthyosaurs gave birth to live young, and the holotype may contain the remains of an embryo in its chest cavity.
History of study
Holotype and naming

A large ichthyosaur skeleton was discovered in the Sasso Caldo site on the Italian side of Monte San Giorgio in 1993. This excavation was part of an operation organized by the Milan Natural History Museum, with the excavation performed by the Gruppo Paleontologico di Besano, a volunteer group. The skeleton, embedded in bituminous level n. 65, was extracted by having the rock layer divided into slabs, each of which were scanned using X-ray analysis. This analysis revealed parts of the skeleton were spread across a total of 33 slabs, which were deposited in the Milan Natural History Museum and given the specimen number BES SC 999. While the slabs are only about 2.7 centimetres (1.1 in) thick, when put together, they cover an area of about 3.5 by 4 metres (11 by 13 ft). The skeleton is flattened from top to bottom, with the exception of the skull, which was instead crushed flat from side to side to a thickness of under 1 centimetre (0.39 in). Most disarticulation of the skeleton is fairly minimal, though the bones of the fingers are scattered. The slabs contain a small mixosaurid in addition to the large ichthyosaur.[2]
By February 1996, 2500 hours of preparation had been performed by four preparators on the specimen, exposing the skull and appendicular skeleton. While the preparation was not yet complete, a preliminary report on this large ichthyosaur by Cristiano Dal Sasso and Giovanni Pinna was published later that year, the authors considering it warranted by the skeleton's distinctiveness and completeness, with the unprepared portions of the skeleton studied through radiography. Dal Sasso and Pinna found the specimen to differ from other shastasaurs, and made it the holotype of a new genus and species, Besanosaurus leptorhynchus. The genus name means "reptile from Besano",[3]: 17 referencing a village in Varese Province, Lombardy, while the Greek words leptòs for "thin" or "slender" and rhynchos for "snout" make up the species name. The authors stated that a more detailed study could be produced once preparation was finished, a task they predicted could take as long as 8000 more hours.[2] The holotype subsequently entered storage as a total of 25 slabs, with a cast of the entire specimen put on display at the same museum. BES SC 999 remains the most complete specimen of Besanosaurus, and is also the oldest definite specimen of a shastasaurid.[1] While Martin Sander and Christiane Faber in 1998 considered it possible that Besanosaurus represented another specimen of the related Shastasaurus,[4] Besanosaurus has otherwise been accepted as a valid taxon.[5][6][7][1]
Further specimens and Mikadocephalus

The holotype of Besanosaurus was not the first shastasaur known from Monte San Giorgio. Two shastasaurid specimens were deposited in the collections of Paläontologisches Institut und Museum der Universität Zürich in the 1920s, both of them being mentioned in passing in the literature of the century. The smaller of the two, numbered PIMUZ T 4376, is a skeleton with a somewhat articulated skull and trunk but disarticulated limbs and tail. While the tail is missing its end the specimen is otherwise nearly complete,[8] and the preservation of the bones is good. The other specimen is cataloged as PIMUZ T 4847 and is very large,[9] and poorly preserved and disarticulated, missing the tail and limbs.[1] Martin Sander and Jean Michel Mazin considered both of these specimens to represent a distinct genera in 1993.[10] The medium-sized specimen was studied by David Cook, who had an abstract published in 1994, in which this specimen was interpreted as pertaining to a new genus.[8] Dal Sasso and Pinna considered this specimen similar to Californosaurus and proportionally distinct from Besanosaurus in their description of the latter genus, following discussion with Robert Appleby.[2] However, Cook's study of PIMUZ T 4376 did not lead to a paper, and the specimen remained incompletely known.[1]

In 1997, Michael Maisch and Andreas Matzke described an ichthyosaur skull housed at the Palaeontological Collection of Tübingen University, cataloged as GPIT 1793/1. This skull is preserved on three shale slabs, and detailed preparation work was done using air abrasion by Fritz Lörcher. While the individual bones are generally fairly well preserved, the skull overall is crushed and strongly disarticulated. Determining it distinct after exhaustive comparisons with other ichthyosaurs, Maisch and Matzke named it the holotype of a new genus and species, Mikadocephalus gracilirostris. The genus name comes from the words Mikado and kephalos (Greek for "head"), referring to the way the skull was preserved, the authors likening its appearance to a game of pick-up sticks, while the species name comes from the Latin words gracilis (slender) and rostrum (snout).[11] However, the authors did not mention Besanosaurus or include it in their comparisons, likely due to them not being aware of its relatively recent publication.[5][1]
Sander argued that Mikadocephalus was probably a junior synonym of Besanosaurus in 2000, noting that they were anatomical similar. Additionally, he found the fact that Besanosaurus and Mikadocephalus would have lived in the same environment to be ecologically implausible given how many other ichthyosaurs had been reported from Monte San Giorgio. He also conisdered it possible that Wimanius, another fragmentary ichthyosaur named by Maisch and Matzke was a juvenile Besanosaurus rather than its own genus, though he also noted that it may instead belong to the same species as the unnamed medium-sized shastasaurid. However, Sander argued that the better-preserved material would need to be studied before definite assignments could be made.[5] Later the same year, Maisch and Matzke defeneded the validity of Mikadocephalus, listing multiple differences between it and Besanosaurus. Additionally, they referred PIMUZ T 4367 to Mikadocephalus gracilirostris, and assigned another specimen to Besanosaurus, PIMUZ T 1895.[6]: 16, 27, 68–69 This specimen consists of a skull associated with postcranial remains, with little limb and tail material. Preparation of this specimen remains incomplete.[1]

In a 2003 book, Christopher McGowan and Motani listed both Mikadocephalus gracilirostris and Wimanius odontopalatus as a species inquirendae, and also considered the high diversity of ichthyosaurs reported from Monte San Giorgio suspicious. They considered both genera potentially synonymous with the much earlier-named Pessosaurus, though remained tentative as the material was not studied firsthand. While Pessosaurus polaris had previously been considered nondiagnostic and thus a nomen dubium, McGowan and Motani suggested that due to its wide historical recognition the name could be revived for some distinctive referred material, noting its similarity to that of the Swiss medium-sized shastasaurid. However, they argued that the medium-sized skeleton, as well as similar Chinese ichthyosaurs, would need to be described in greater detail before they could assess the validity of P. polaris.[7]: 127–128 Maisch argued that Wimanius and Mikadocephalus were distinct and valid in 2010, stating that there were was no morphological or phylogenetic support for their synonymy, and kept Besanosaurus and Mikadocephalus separate as well.[12] Otherwise, however, little further research was published on the taxonomy of Besanosaurus.[1]
In 2021, a paper by Gabriele Bindellini and colleagues detailing the skull anatomy of Besanosaurus was published. The authors studied six shastasaurid specimens from Monte San Giorgio, namely the holotypes of Besanosaurus and Mikadocephalus, PIMUZ T 4376, PIMUZ T 4847, PIMUZ T 1895, and BES SC 1016. This last specimen is an incomplete, somewhat flattened, partially articulated skull preserved in dolomite, which was analyzed with CT scans. Most of the specimens had not previously had their skull anatomy described in detail, and BES SC 1016 had not been studied before. The authors reassessed the status of Mikadocephalus, finding many of the features initially used to distinguish it to be present in Besanosaurus as well, and were not able to find any distinguishing characteristics between the two genera. Therefore, they synonymized Mikadocephalus gracilirostris with Besanosaurus leptorhynchus, and referred the other four specimens to the latter species as well, since they also showed identifying features of Besanosaurus.[1]
Description

Ichthyosaurs like Besanosaurus were marine reptiles, with flippers for limbs and a fin on the tail.[5][13] While later ichthyosaurs developed fish-shaped body plans, Besanosaurus is more elongate,[14][13] and resembles Cymbospondylus in overall shape. The skull of Besanosaurus is proportionately quite small, making up less than 10% of the animal's total length in the holotype. In addition to having an elongate trunk, about half of the total length of Besanosaurus is formed by its very long tail.[2] The holotype specimen measures 5.065 meters (16.62 ft) long from its snout tip to tail tip. The specimen PIMUZ T 4847 was estimated by Bindellini and colleagues in 2021 to have a total length of 8 meters (26 ft), and is the largest known specimen of Besanosaurus, indicating that Besanosaurus is the largest known ichthyosaur in its environment.[1]
Skull
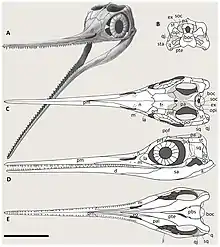
About two-thirds of the skull of Besanosaurus is occupied by its very long, thin snout lined with small teeth. This distinctive snout is strongly demarcated from the rear part of the skull by a constriction.[2][1] Well-preserved Besanosaurus skulls show drawn-out slotlike external nares, openings which housed the nostrils in life. Behind the external nares are the elliptical orbits, somewhat longer than tall. Within the orbits are the sclerotic rings, circles of bony, quadrangular plates that supported the eyeballs. Bindellini and colleagues estimated a total of between 15 and 17 of these plates per ring in Besanosaurus. While large relative to the orbits, the rings are smaller relative to the animal's overall length than typical for parvipelvian ichthyosaurs. Further back, the top of the skull is perforated by another pair of openings, the supratemporal fenestrae, which are rather small in Besanosaurus. Extending forwards from each supratemporal fenestra is a depression, forming roughly elliptical terraces in front of the fenestrae. The region behind the orbits is short, occupying 13% of the skull's length,[2] though it is still long compared to some parvipelvians.[1]
In the upper jaw, the snout is formed by the elongation of the tooth-bearing premaxillae. The front ends of these bones are blunt, while their rear ends are forked, with processes extending both above and below the external nares. The back edges of the external nares are formed by the upwards-directed processes of the maxillae, gracile, triradiate bones behind the premaxillae.[1] While initially interpretted as not forming part of the narial border in the holotype,[2] further specimens revealed that the nasals, which run along the top of the snout, do participate in the upper back corners of the external nares. Along the midline of the skull, between the front ends of the temporal fenestrae, is a small opening called the pineal foramen, whose border is formed by the frontals in front and parietals on the sides and back. A sagittal crest also runs along these two pairs of bones. The frontals are long, broad,[2] and flat bones involved in the supratemporal terraces, though they do not form any part of the borders of the supratemporal fenestrae. The parietals bear a variety of prominent surfaces for jaw muscle attachment, including the saggital crest and their contributions to the rims of the supratemporal fenestrae.[1]
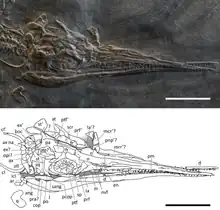
The front portions of the orbits are formed by a pair of crescentic bones called lacrimals. The prefrontals and postfrontals make up the top of the orbital rim, where they are thickened. The prefrontals also extend forwards, where they contact the ascending processes of the maxillae. The back edges of the orbits are formed by the postorbitals, which also reach upwards and enter the supratemporal fenestrae. On the lower regions of the sides of the skull, wide portions of the postorbitals are visible, which Bindellini and colleagues considered potentially autapomorphic of the genus. The jugals are thin, strip-like bones bent at right angles. The articulations with other bones holding it in place were rather weak, and resultingly the jugals were often disarticulated from the skull during fossilization. The squamosals of Besanosaurus are large and quadrangular, as is typical for ichthyosaurs with longer cheek regions, and seen in many other shastasaurids.[1]
The jaw joint was formed by the robust, kidney-shaped quadrates in the cranium. Each quadrate has a triangular eminence on the lower part of its inner margin. In 1997, Maisch and Matzke considered this to be a unique feature of Mikadocephalus, as at the time it was only known in the holotype skull of that taxon;[11] however, Bindellini and colleagues identified this on multiple Besanosaurus specimens in 2021, as well as a specimen of Guanlingsaurus, and therefore suggested that it may be a fairly widespread feature of shastasaurids.[1] The vertebral column articulated with a convex eminence on the back of the skull known as the occipital condyle.[1][2] Above this, the spinal cord entered through an opening called the foramen magnum, which Bindellini and colleagues reconstructed as rounded and roughly pentagonal in Besanosaurus.[1] The stapedes of Besanosaurus, a pair of bones in the braincase, are broad-ended but slender-shafted.[1][2] The openings between the pterygoids in the rear part of the palate was interpretted as wide by Maisch and Matzke in their description and reconstruction of GPIT 1793/1;[11][15] however, Bindellini and colleagues in their redescription argued that this was not necessarily the case due to the specimen's disarticulation, and noted that the width of the interpterygoid vacuity was of typical width for a shastasaurid in BES SC 1016.[1] The pterygoids possess long processes for articulation with the quadrates.[11][1]
Most of the lower jaw is formed by the long, thin toothbearing dentaries. Behind the dentaries, the large surangulars form the top portion of the rear part of the lower jaw. The upper margin of each surangular bears two eminences, a taller, pointed one immediately in front of the jaw joint and another, lower one further forwards. Based on comparisons with Chaohusaurus, Bindellini and colleagues interpretted the taller of the two projections as the coronoid process, formed through the fusion of the coronoid to the surangular. The coronoid process is massive in Besanosaurus, only rivaled in its development by that of Phantomosaurus. The teeth of Besanosaurus are all similar in shape, being small, cone-shaped, and acutely pointed. The tooth crowns are lined with vertical ridges, though lack cutting edges, while the tooth roots bear even stronger ridging. The teeth are separated from each other by a rather wide amount of space and interlock when the jaw is closed. The frontmost teeth in the upper jaw are the longest, and somewhat curved, while the back teeth are shorter and thicker, with a similar pattern of size and shape also present in the lower jaw. How the teeth are implanted varies through the tooth row; throughout most of the upper jaw, the teeth are set into sockets, with the exception of the rear 30% of the maxillary teeth,[11] which are implanted in a groove. Conversely, in the lower jaw, the front teeth are located in a groove while the back teeth are set into sockets.[1]
Vertebrae and ribs

Besanosaurus has 60 vertebrae located in front of its hips (presacrals); 11 of these are neck vertebrae, located in front of the shoulders. Behind the presacral vertebrae, there are 2 vertebrae in the hip region and 139 in the tail. The neck vertebrae have prominent zygapophyses (bony projections of the vertebrae) and large, robust neural spines with roughly round cross-sections. The roughly rectangular neural spines of the trunk vertebrae are also tall, and are more than half as wide from side to side as they are from front to back. The vertebral bodies, known as centra, of the trunk vertebrae are at most half as long as tall. The front and back faces of all the centra are concave, being bowed inwards to a very thin layer of bone if not an opening.[2] The articular surfaces for the ribs on the sides centra do not extend all the way forwards to their front edges.[16] The frontmost cervical ribs (those attached to the neck vertebrae) are double-headed, unlike the other, single-headed cervical ribs behind them. The dorsal (trunk) ribs bear grooves along their shafts. The heavily built gastralia (belly ribs) are united along the animal's midline.[2]
Most of the neural spines of the tail vertebrae slope backwards, except near its end, where they lean in the opposite direction. Dal Sasso and Pinna were not able to determine if Besanosaurus had any wedge-shaped caudal centra due to how the tail was preserved. Such vertebrae would form a downward kink known as a tailbend. However, Dal Sasso and Pinna compared the vertebral shape with that of Cymbospondylus and suggested that the tail of Besanosaurus would have been fairly straight.[2] However, Bindellini and colleagues reported a tailbend in PIMUZ T 1895.[1] The articular faces of the centra of the tail vertebrae vary in shape; the front ones are roughly triangular, the middle are hexagonal, and the rear are elliptical. Beneath the tail vertebrae are the chevrons, Y-shaped bones which are not very long in Besanosaurus.[2]
Appendicular skeleton
.jpg.webp)
The sickle-shaped scapulae of Besanosaurus are wide, with broadly expanded upper portions. Their front lower edges are short and mildly bowed inwards. Another pair of shoulder bones, the coracoids are shaped like axe heads, with their upper margins heavily expanded and their lower ends fan-shaped. In Cymbospondylus, each coracoid is pierced by a foramen, however, no such openings are present in those of Besanosaurus. The clavicles (collarbones) of Besanosaurus are thin and broadest at their middles, and the interclavicle, a midline bone located between them, does not appear to have been present. The forelimbs of Besanosaurus are about 15% longer than its hindlimbs. Each flipper bears a total of four digits, three of them well-developed and a fourth being an accessory digit (a digit not originating from the wrist or ankle). The humeri are very short, being wider than they are long, and have a rounded profile. Both the front and back edges of the humeri are bowed inwards, though this emargination is more notch-like on the front edges.[2] The lower arm bones as wide as they are long, with the radii being the larger of the two pairs. The roughly quadrangular radii have constricted middles, while the ulnae behind them are rounded and somewhat smaller.[2][17] The carpals, metacarpals, and phalanges (finger bones) are also rounded, the latter two types being having the shape of circles.[2]
The upper hip bones of Besanosaurus, the ilia, are rather wide, with their upper ends enlarged and directed inwards and their lower ends more heavily expanded. The pubic bones (front lower hip bones) are vaguely circular in shape. They each bear a well-developed but narrow notch extending to the edge of the bone, rather than an enclosed foramen.[2] The femora are longer than wide, though only by a by a factor of 1.22 and are resultingly still quite stout. The femora are narrowest midshaft, broadening at their upper ends. The tibiae and fibulae are also constricted at their middles and narrower than the radii and ulnae. The tibiae are the longer of the two pairs of lower leg bones, and the fibulae have heavily expanded lower ends. Compared with the corresponding elements of the forelimb, the tarsals, metatarsals, and phalanges of the hindflippers are less robust. Furthermore, while round, the phalanges of the pes are oblong and constricted.[2]
Classification

While Besanosaurus has generally been considered a shastasaurid,[18] how it is related to other shastasaurids is unclear due to the poorly understood phylogeny of the group.[1] To determine the phylogenetic affinities of Besanosaurus, Dal Sasso and Pinna coded it into a data matrix created that was created by Jack Callaway in 1986. They found shastasaurids to be divisible into two main groups, termed Cymbospondylinae and Shastasaurinae by Callaway, with the former being a grade leading up to the latter.[2][6]: 4–5 Besanosaurus was placed in an intermediate position between these two subfamilies by the analysis. While Dal Sasso and Pinna noted that Besanosaurus superficially resembled Cymbospondylus in overall body shape, they found it to compare more favorably to the shastasaurines in the fine points of its anatomy, and thus tentatively assigned it to Shastasaurinae.[2] The phylogenetic analysis of Dal Sasso and Pinna was one of the earlier of such studies on ichthyosaurs, and subsequent publications have criticized it for a variety of reasons. Key among these issues is that their analysis did not test their assumption that Shastasauridae was a natural group, as their matrix did not include any non-shastasaurid ichthyopterygians other than Grippia. The usage of Cymbospondylinae for a paraphyletic group and the inclusion of highly fragmentary taxa was also criticized.[6]: 4–5 [7]: 144
The first comprehensive, in-depth study of overall ichthyosaur relationships was done by Motani in 1999.[12][7]: 144 The traditional concept of Shastasauridae was not recovered by his analysis, with some "shastasaurids" such as Cymbospondylus representing earlier branches of ichthyosaur evolution while others like Californosaurus were found to be more closely related to the later, post-Triassic ichthyosaurs. Some shastasaurids, however, were found to form a natural group in his analysis, Besanosaurus among them. Based on the results of his analysis, he placed Besanosaurus in Shastasauridae, but outside Shastasaurinae, formed by Shastasaurus and Shonisaurus. Merriamosauria was named for the group including the shastasaurids and later ichthyosaurs. Motani noted that some parts of the phylogeny were unstable, and that many traditional shastasaurids were "problematic" due to their incompleteness.[19] Another large phylogenetic study conducted by Sander was published the following year,[18] focusing on the more completely known ichthyosaurs. Unlike Motani, Sander found the three shastasaurids he included in his analysis to form a natural group, with Besanosaurus being the sister taxon of Cymbospondylus and Shonisaurus being basal to the two.[5] This analysis was criticized by subsequent work for only using a small sample of ichthyosaurs and encoding proportional characters based on arbitrary divisions. Concerns were also raised about the reproducibility of his results and the extremely high amount of homoplasy (convergent evolution of features by unrelated groups) implied by his cladogram.[12][7]: 145–146
A third major analysis of ichthyosaur relationships was done by Maisch and Matzke later in 2000, featuring many similarities to Motani's analysis but based on a larger data set.[6][12] Like Motani, Maisch and Matzke found the traditional shastasaurids to not be a natural group, however, they also found Motani's concept of Shastasauridae to be a grade leading up to the post-Triassic ichthyosaurs and their relatives, with Besanosaurus as the first of these taxa to branch off. As it was considered separate at the time, Mikadocephalus was also coded into their analysis, and found to be more closely related to the post-Triassic ichthyosaurs than Besanosaurus.[12][6]: 96–97, 100 They had previously found a similar result in a smaller study they conducted in 1997, which found Mikadocephalus to be more closely related to the post-Triassic ichthyosaurs.[11][12] However, the phylogeny of Maisch and Matzke (2000) has subsequently been criticized for using a hypothetical ancestor, rather than actual non-ichthyosaurian taxa, as an outgroup.[17][7]: 146 McGowan and Motani considered Besanosaurus to fall outside of Merriamosauria in 2003, and named a new monotypic family, Besanosauridae, to contain it.[7]: 70 Maisch also used this family for Besanosaurus in 2010, though he considered it to be within Merriamosauria. He classified Mikadocephalus in a separate family, Guanlingsauridae, alongside Guanlingsaurus in the same paper.[12]

In 2011, Philippa Thorne and colleagues ran an analysis based on an updated version of Motani's 1999 data matrix.[20] Their analysis found Besanosaurus to be part of the shastasaurid clade (natural group). The results of another comprehensive analysis by Cheng Ji and colleagues were published in 2016, and was based on a much larger data set than previous studies. A large, well-supported clade of shastasaurids was recovered by their study, with Besanosaurus as the earliest diverging member.[17] Another series of very large analyses were run by Benjamin Moon in 2017, created by revising and compiling previous analyses. The analyses, however, were unable to clearly elucidate the relationships of many early merriamosaurs, making it unclear whether or not Besanosaurus was a shastasaur.[18] In their 2021 study, Bindellini and colleagues updated the codings for Besanosaurus and some other shastasaurids in the analysis of Ji and colleagues, preferring this study over Moon's larger one as it was constructed with more direct observation of the specimens. They found multiple different configurations of shastasaurids, with the group either being a grade or a clade, with the former arrangement found with marginally more frequency. Within the grade, Besanosaurus was found to be the earliest diverging member, while within the clade it was either in a similar position or in a smaller nested group with Guizhouichthyosaurus and "Callawayia" wolonggangense. Due to this instability, Bindellini and colleagues considered shastasaurid relationships to still be ambiguous.[1]
The following cladograms depict the three hypotheses recovered by the analysis of Bindellini and colleagues, 2021.[1]
|
Cladogram showing paraphyletic Shastasauridae:
|
Cladogram showing monophyletic Shastasauridae:
|
Cladogram showing monophyletic Shastasauridae:
|
Paleobiology

Ichthyosaurs like Besanosaurus were very well-adapted to their marine existence, spending their entire lives in the water and showing reduced levels of ossification of their bones. Nevertheless, ichthyosaurs still would have breathed air. Ichthyosaurs were active animals with high metabolic rates, able to maintain their body temperatures.[5][21][22] Intermediate-grade ichthyosaurs likely used anguilliform (eel-like) locomotion, swimming by undulating their entire body from side to side, owing to their long tails without strong bends and their long, flexible trunks. This differs from the more tail-driven locomotion presumed for the more spindle-shaped parvipelvians.[23][14][5] While not used in propulsion, ichthyosaur limbs would have been used to steer and keep the body upright.[5] The large eyes of ichthyosaurs indicate that vision would have been an important sense for them.[5][7]: 148 Besanosaurus is the earliest known long-snouted diapsid that attained a large body size. Bindellini listed many potential advantages of large size, including it allowing various feeding techniques, being an anti-predator adaptation, and resulting in more efficient thermoregulation.[1] A 2020 survey of ichthyosaurs from Monte San Giorgio by Judith Pardo-Pérez and colleagues found that shastasaurids like Besanosaurus were more likely to show skeletal injuries than the smaller mixosaurids, but showed less frequent injuries than Cymbospondylus, thought to have preyed upon larger animals. The holotype of Besanosaurus exhibits ankylosed (pathologically fused) metatarsals.[14]
Diet and feeding

.jpg.webp)
Dal Sasso and Pinna considered Besanosaurus to likely have been specialized on feeding on coleoid cephalopods, based on the shape and small size of its teeth.[2] Bindellini and colleagues elaborated on the tooth anatomy of Besanosaurus, noting that the slender teeth in front would have been suited for piercing small prey while the slightly blunter ones further back suited for catching cephalopods by grasping. Direct evidence of Besanosaurus feeding on cephalopods is present in the form of a Phragmoteuthis-like hooklet preserved as stomach contents in the holotype.[1] However, following the discovery of a specimen of Guizhouichthyosaurus, a supposed cephalopod predator, with a considerable portion of a large thalattosaur in its stomach region, Da-Yong Jiang and colleagues speculated in 2020 that other large ichthyosaurs adapted for consuming cephalopods may have been able to take on large prey. The researchers noted that despite the rather delicate construction of the skull of Besanosaurus, a Mixosaurus would have been able to fit inside its mouth.[24] However, Bindellini and colleagues doubted that Besanosaurus would have been a predator of large animals, citing its long, slender snout, arguing that a diet of small animals was more likely.[1]
The large coronoid processes and rough, concave outer faces of the surangulars would have anchored strong jaw musculature. While the thin jaws of Besanosaurus would not have been able to support a powerful bite, Bindellini and colleagues noted that it could instead have allowed the jaws to rapidly close. The length of the jaws would have caused their tips to move quite quickly during biting. Additionally, the long, thin snout could have been swung efficiently through the water, allowing quick up and down or side to side movements. Altogether, this configuration is ideal for feeding on prey items that are small and agile. Long, thin snouts have convergently evolved multiple times in aquatic amniotes with similar feeding habits. As modern long-snouted predators such as some dolphins are piscivorous, Bindellini and colleagues suggested that Besanosaurus may have consumed fish as well as coleoids. Bindellini and colleagues also noted that the large size of Besanosaurus could have helped with prey capture as well by providing more inertia, with the head being able to have broad movement while the body stayed in place. The authors considered the animal's large size to have potentially resulted from its foraging method being particularly efficient.[1]
Reproduction and growth

Dal Sasso and Pinna discovered three or four small series of vertebrae within the holotype's chest through radiography. They interpretted the remains as pertaining to embryos, noting that a placement so far forwards in the body was not unheard of among ichthyosaurs. They also considered it doubtful that Besanosaurus would have been able to take on mixosaurid-sized prey.[2] In a 2021 abstract, Feiko Miedema and colleagues regarded the embryonic remains as pertaining to one embryo.[25] In 2023, Miedema and colleagues interpretted the embryo as being oriented so that it would have been born tail first, as typical in merriamosaurs. Historically, this orientation was thought to have evolved in ichthyosaurs to reduce the likelihood of the newborns drowning during birth; however, after surveying different vivaporous amniotes, Miedema and colleagues argued that the evidence for such an adaptation was lacking, and instead proposed that birth orientation was related to which way was easier to push the fetus through the birth canal or which way made affected the mobility of the pregnant adult less.[26] However, embryos are not the only interpretation of these remains. Jiang and colleagues in 2020 were doubtful about the inability of Besanosaurus to have consumed mixosaurids, and argued that it could not be ruled out that the supposed embryos may be stomach contents instead, noting that the vertebrae were the right size for an immature mixosaurid, and were more ossified than would be expected in an embryo.[24]
As ichthyosaurs grow, their skulls typically become proportionately smaller. In PIMUZ T 4376, the skull is roughly half the length of the trunk; whereas it is only about a third as long as the trunk in the holotype.[1] This difference was considered too extreme to be the result of ontogeney by Maisch and Matzke in 2000, arguing the specimens were too similar in size. Thus, they used this feature to distinguish Mikadocephalus from Besanosaurus.[6]: 34–25 Bindellini and colleagues, however, did not find ontogeny to be an unreasonable explanation for this proportional difference in their 2021 study, instead finding the six specimens they studied to fit into a growth series when ordered by size. Unlike the skull length-body length ratio of Besanosaurus, the researchers found the orbit size and mandible length to increase at the same rate, and the tooth anatomy of Besanosaurus to not change with ontogeny.[1]
References
- Bindellini, G.; Wolniewicz, A.S.; Miedema, F.; Scheyer, T.M.; Dal Sasso, C. (2021). "Cranial anatomy of Besanosaurus leptorhynchus Dal Sasso & Pinna, 1996 (Reptilia: Ichthyosauria) from the Middle Triassic Besano Formation of Monte San Giorgio, Italy/Switzerland: taxonomic and palaeobiological implications". PeerJ. 9: e11179. doi:10.7717/peerj.11179. PMC 8106916. PMID 33996277.
- Dal Sasso, C.; Pinna, G. (1996). "Besanosaurus leptorhynchus n. gen. n. sp., a new shastasaurid ichthyosaur from the Middle Triassic of Besano (Lombardy, N. Italy)". Paleontologia Lombarda. 4: 3–23.
- Dal Sasso, C. (2004). Dinosaurs of Italy. Bloomington, Indiana: Indiana University Press. ISBN 978-0-253-34514-1.
- Sander, P.M.; Faber, C. (1998). "New finds of Omphalosaurus and a review of Triassic ichthyosaur paleobiogeography". Paläontologische Zeitschrift. 72 (1): 149–162. doi:10.1007/BF02987823.
- Sander, P. M. (2000). "Ichthyosauria: their diversity, distribution, and phylogeny". Paläontologische Zeitschrift. 74 (1): 1–35. doi:10.1007/BF02987949. S2CID 85352593.
- Maisch, M. W.; Matzke, A. T. (2000). "The Ichthyosauria". Stuttgarter Beiträge zur Naturkunde, Serie B. 298: 1–159.
- McGowan, C.; Motani, R. (2003). Sues, H.D. (ed.). Handbook of Paleoherpetology Part 8: Ichthyopterygia. Munich: Verlag Dr. Friedrich Pfeil. ISBN 3-89937-007-4.
- Cook, D.H.N. (1994). "A new ichthyosaur genus from the Middle Triassic of Monte San Giorgio, Switzerland". Journal of Vertebrate Paleontology. 14 (3): 21A–22A.
- Sander, P.M. (1989). "The large ichthyosaur Cymbospondylus buchseri, sp. nov., from the Middle Triassic of Monte San Giorgio (Switzerland), with a survey of the genus in Europe". Journal of Vertebrate Paleontology. 9 (2): 163–173.
- Sander, P.M.; Mazin, J.M. (1993). "The paleobiogeography of Middle Triassic ichthyosaurs: The five major faunas". Paleontologia Lombarda, Nuova Serie. 2: 145–152.
- Maisch, M.W.; Matzke, A.T. (1997). "Mikadocephalus gracilirostris n. gen., n. sp., a new ichthyosaur from the Grenzbitumenzone (Anisian-Ladinian) of Monte San Giorgio (Switzerland)". Paläontologische Zeitschrift. 71 (3): 267–289. doi:10.1007/BF02988496.
- Maisch, M.W. (2010). "Phylogeny, systematics, and origin of the Ichthyosauria – the state of the art" (PDF). Palaeodiversity. 3: 151–214.
- Motani, R. (2005). "Evolution of fish-shaped reptiles (Reptilia: Ichthyopterygia) in their physical environments and constraints". Annual Review of Earth and Planetary Sciences. 33: 395–420. Bibcode:2005AREPS..33..395M. doi:10.1146/annurev.earth.33.092203.122707.
- Pardo-Pérez, J.M.; Kear, B.P.; Maxwell, E.E. (2020). "Skeletal pathologies track body plan evolution in ichthyosaurs". Scientific Reports. 10 (1): 4206. Bibcode:2020NatSR..10.4206P. doi:10.1038/s41598-020-61070-7. PMC 7060314. PMID 32144303.
- Maisch, M.W.; Matzke, A.T. (1999). "Observations on Triassic ichthyosaurs. Part V. The skulls of Mikadocephalus and Wimanius reconstructed". Neues Jahrbuch für Geologie und Paläontologie, Monatshefte. 1999 (6): 345–356. doi:10.1127/njgpm/1999/1999/345.
- Dal Sasso, C.; Insacco, G.; Chiarenza, A.A.; Di Franco, D.; Reitano, A. (2014). "First record of ichthyosaurs in Sicily (Upper Triassic of Monte Scalpello, Catania Province)". Rivista Italiana di Paleontologia e Stratigrafia. 120 (1): 71–82.
- Ji, C.; Jiang, D. Y.; Motani, R.; Rieppel, O.; Hao, W. C.; Sun, Z. Y. (2016). "Phylogeny of the Ichthyopterygia incorporating recent discoveries from South China". Journal of Vertebrate Paleontology. 36 (1): e1025956. doi:10.1080/02724634.2015.1025956. S2CID 85621052.
- Moon, B.C. (2017). "A new phylogeny of ichthyosaurs (Reptilia: Diapsida)" (PDF). Journal of Systematic Palaeontology. 17 (2): 1–27. doi:10.1080/14772019.2017.1394922. S2CID 90912678. Archived from the original on 2020-03-18. Retrieved 2023-09-30.
{{cite journal}}: CS1 maint: bot: original URL status unknown (link) - Motani, R. (1999). "Phylogeny of the Ichthyopterygia" (PDF). Journal of Vertebrate Paleontology. 19 (3): 473–496. Bibcode:1999JVPal..19..473M. doi:10.1080/02724634.1999.10011160. Archived from the original (PDF) on 2012-04-15.
- Thorne, P.M.; Ruta, M.; Benton, M.J. (2011). "Resetting the evolution of marine reptiles at the Triassic-Jurassic boundary". Proceedings of the National Academy of Sciences. 108 (20): 8339–8344. doi:10.1073/pnas.1018959108.
- Naish, D. (2023). "Shark-shaped reptiles: The ichthyosaurs and their kin". Ancient Sea Reptiles: Plesiosaurs, Ichthyosaurs, Mosasaurs, and More. Washington, DC: Smithsonian Books. pp. 94–123. ISBN 978-1-58834-727-5.
- Nakajima, Y.; Houssaye, A.; Endo, H. (2014). "Osteohistology of the Early Triassic ichthyopterygian reptile Utatsusaurus hataii: Implications for early ichthyosaur biology". Acta Palaeontologica Polonica. 59 (2): 343–352. doi:10.4202/app.2012.0045.
- Gutarra, S.; Moon, B. C.; Rahman, I. A.; Palmer, C.; Lautenschlager, S.; Brimacombe, A. J.; Benton, M. J. (2019). "Effects of body plan evolution on the hydrodynamic drag and energy requirements of swimming in ichthyosaurs". Proceedings of the Royal Society B. 286 (1898): 20182786. doi:10.1098/rspb.2018.2786. PMC 6458325. PMID 30836867.
- Jiang, D.Y.; Motani, R.; Tintori, A.; Rieppel, O.; Ji, C.; Zhou, M.; Wang, X.; Lu, H.; Li, Z.G. (2020). "Evidence supporting predation of 4-m marine reptile by Triassic megapredator". iScience. 23 (9): 101347. doi:10.1016/j.isci.2020.101347. PMC 7520894. PMID 32822565.
- Miedema, F.; Bindellini, G.; Dal Sasso, C.; Scheyer, T.M.; Maxwell, E.E. (2021). "Conserved cranial development and early ontogeny in Triassic and Jurassic ichthyosaurs". European Association of Vertebrate Palaeontologists 18th Annual Meeting: 116.
- Miedema, F.; Klein, N.; Blackburn, D.G.; Sander, P.M.; Maxwell, E.E.; Griebeler, E.M.; Scheyer, T.M. (2023). "Heads or tails first? Evolution of fetal orientation in ichthyosaurs, with a scrutiny of the prevailing hypothesis". BMC Ecology and Evolution. 23 (12). doi:10.1186/s12862-023-02110-4.
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||||